

Research Article | DOI: https://doi.org/10.31579/2834-5029/021
Steroid Hormones
- Rehan Haider *
Riggs pharmaceutical, Department of Pharmacy, University of Karachi, Pakistan.
*Corresponding Author: Rehan Haider, Riggs pharmaceutical, Department of Pharmacy, University of Karachi, Pakistan.
Citation: Rehan Haider, (2023), Steroid Hormones, International Journal of Biomed Research. 2(3): DOI: 10.31579/2834-8087/021
Copyright: © 2023, Rehan Haider. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 05 May 2023 | Accepted: 26 May 2023 | Published: 02 June 2023
Keywords: estrogens; hormones; neuro protection; neurosteroids; neurotransmitters; ovarian continuum; progesterone
Abstract
Steroids are lipophilic molecules that are used as chemical messengers by organisms ranging in complexity from water molds to humans. In vertebrates, steroids act on a large range of tissues and influence many elements of biology which consist of sexual differentiation, reproductive body structure, osmoregulation, and intermediate metabolism. important websites of steroid synthesis and secretion include the ovaries, testes, adrenals, and placenta. primarily based totally on the space of a target internet site online from the internet site of synthesis and secretion, steroid hormones can be labeled as each endocrine (faraway goal tissue), paracrine (neighboring cells), or autocrine (same cell) elements. while secreted into the surroundings, steroid scan scans also act as pheromones with the aid of conveying facts to other organisms. thanks to the pervasive effects of steroids in vertebrate biology, several pathologic states can occur due to issues associated with steroid hormone action. These ailment states encompass most cancers, steroid insensitivity, and steroid synthesis.
The cause for this financial disaster is to provide a top-level view of steroid synthesis, steroid hormone results in ordinary frame shape, molecular and biochemical mechanisms of action of steroid hormones, and pathologic states associated with steroid hormone movement.
Introduction
Steroid Hormone Synthesis
Steroid hormones are lipid molecules derived from a common cholesterol precursor, Cholestane (C27). There are four major classes of steroid hormones, progestins, androgens, estrogens, and corticoids, which contain 21, 19, 18, and 21 carbons, respectively. Steroid hormones are synthesized by dehydrogenases and cytochrome P450 enzymes, which catalyze hydroxylation and dihydroxylation-oxidation reactions. Eukaryotic cytochromes P450 are membrane-bound enzymes expressed in either the inner mitochondrial or endoplasmic reticulum membranes of steroid-synthesizing tissues. A common and important rate-limiting step for the synthesis of all steroid hormones is the cleavage of the side chain from cholesterol (C27) to yield pregnenolone (C21), the common branch point for the synthesis of progestins, corticoids, androgens, and, hence, estrogens (Figure. 1). Expression of the side-chain cleavage enzyme cytochrome P450scc (cytP450scc), which converts cholesterol to pregnenolone, is one of the unique features of steroidogenic cells that participate in de novo steroid synthesis. In vertebrates, the synthesis and secretion of gonadal and adrenal steroid hormones are regulated by tropic hormones from the anterior pituitary such as follicle-stimulating hormone (FSH), luteinizing hormone (LH), and adrenocorticotropic hormone (ACTH). Mineralocorticoid are also regulated by ion concentrations and Circulating angiotensin II levels common regulatory mechanisms for steroid synthesis and launch are bad feedback loops in which multiplied circulating stages of steroids suppress the production of tropic hormones by way of acting on particular sites inside the brain and anterior pituitary. The complicated interplay amongst special additives of the hypothalamic-pituitary-gonad (HPG)/adrenal (HPA) axes is a vital function of the endocrine physiology.
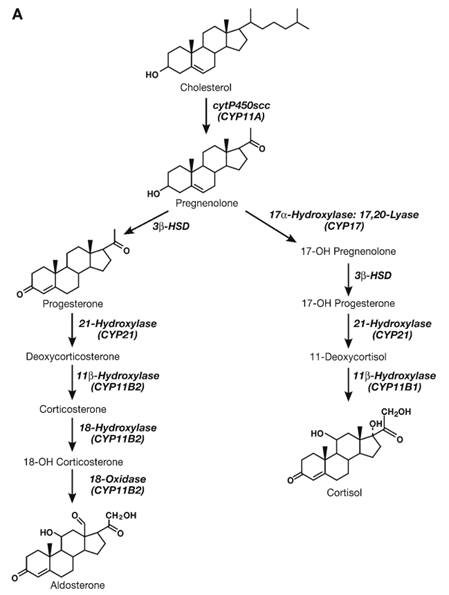
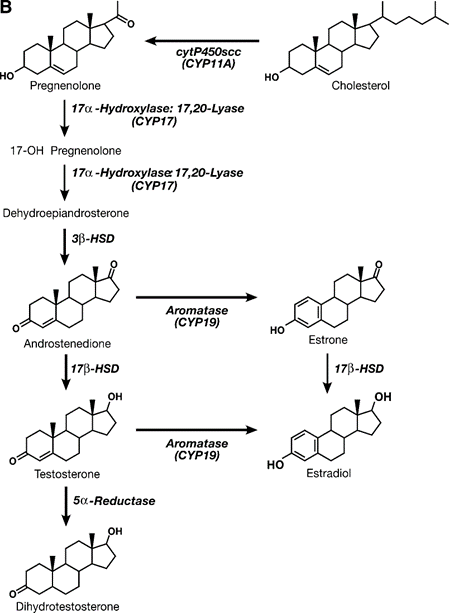
Synthesis of Progesterone
Pregnenolone serves as a principal precursor to all the other steroid hormones synthesized by using the ovary, testes, or adrenals. It seems that the rate-limiting step for the synthesis of progesterone is facet-chain cleavage of LDL cholesterol by way of P450scc. Pregnenolone is then converted into progesterone through threeβ-hydroxysteroid dehydrogenase (3β-HSD). thus, deficiencies in both. P450scc and 3β-HSD have profound outcomes on the synthesis of all steroids. In the ovary, progesterone is produced at all tiers of follicular development as an intermediate for androgen and estrogen synthesis, but turns into a primary secretory product throughout the peri- and post-ovulatory (luteal) stages. Progesterone synthesis is controlled by FSH during the early stages of follicular genesis and, following the purchase of LH receptors, becomes sensitive to LH later in the ovarian cycle. The synthesis. of progesterone with the aid of the corpus luteum is stimulated at some point in early pregnancy by growing levels of chorionic gonadotropin. similarly, the placenta secretes excessive stages of progesterone at some point in pregnancy, although a distinct isozyme of 3β-HSD is involved in the synthesis.
Synthesis of Androgen
Androgen is typically synthesized and secreted with the aid of the Leydig cells of the testes, thecal cells of the ovary, and cells within the reticular location of the adrenals. In most tetrapod vertebrates, testosterone is the dominant circulating androgen. Testicular synthesis and secretion of testosterone are inspired by the use of circulating LH, which upregulates 17? -hydroxylase: C-17,20- lyase, a price-proscribing enzyme for the conversion of C21 to C19 steroids. Once taken up using intentional tissues, testosterone may be reduced with a useful resource of 5? -reductase to yield an additional lively androgen metabolite 5.-dihydrotestosterone (5? -DHT). Testosterone and androstenedione can also be converted into estrogens together with 17.-estradiol (E2) or estrone, through aromatization. Aromatization has finished the usage of a cytochromeP450 aromatase enzyme that is expressed in the granulosa cells of the ovary, Leydig cells of the testes, and many incredible tissues that comprise the placenta, mind, pituitary gland, liver, and adipose tissue. Sincerely, are some of the outcomes of circulating testosterone due to conversion into every 5? -DHT or E2 internal-purpose tissues?
Synthesis of Estrogen
Estrogen and progestins are synthesized and secreted commonly through maturing follicles, corpora lutea of ovaries, and the placenta sooner or later in pregnancy. The maximum amount of estrogen secreted is E2, and the critical progestin is progesterone. The profile of the synthesis of estrogen changes throughout follicular genesis at some unspecified time in the future. Underneath the effect of LH, the thecal cells synthesize and secrete androstenedione and testosterone, which diffuse at some stage in the basement membrane and are in the long run aromatized to estrone and E2, respectively, via the use of granulosa cells. The volume of aromatase and, in the end, estrogens produced within the granulosa cells are below the management of FSH within the course of mid-follicular ranges. Later inside the cycle, the follicle/corpora lutea have unique numbers of LH receptors, and LH starts to evolve to alter E2 manufacturing. Throughout pregnancy, the placenta uses androgen precursors from the fetal adrenal gland and secretes large amounts of E2. Similarly, in male vertebrates, many target tissues, together with pituitary cells and hypothalamic neurons, convert circulating testosterone to E2.
Synthesis of Corticoid
Corticoids are divided into glucose- and mineralocorticoid. The foremost human glucocorticoid, cortisol, is synthesized within the zona fasciculata of the adrenal cortex. The synthesis of cortisol was initiated hydroxylation of progesterone at positions 17α, 21 (CYP21), and 11β (CYP11B1). The synthesis of cortisol.is beneath the management of an anterior pituitary hormone, ACTH, and a terrible feedback mechanism where in expanded cortisol suppresses the discharge of ACTH The dominant human mineralocorticoid is aldosterone, which is produced inside the zona glomerulosa of the adrenal. The synthesis of aldosterone includes corticosterone synthesis, followed by hydroxylation and oxidation at C18 to yield aldosterone. The synthesis of aldosterone is regulated directly via stages of potassium, serum-binding, and circuitously with the resource of the effects of sodium degrees and blood volume on degrees of angiotensin II
Serum-Binding Proteins
After synthesis, steroids are transported to their target tissues via the bloodstream. The hydrophobic nature of steroid hormones consequences in low solubility in water; therefore, transport proteins, recognized as s proteins, help in the transport of steroid hormones to their target tissues. This cargo is done. Binding of steroid hormones to particular high-affinity ligand-binding areas (LBD) in serum-binding proteins. Five serum-binding proteins. were diagnosed with corticosteroid-binding globulin, retinol-binding protein, sex hormone-binding globulin (SHBG), thyroxine-binding globulin, and vitamin D-binding protein. As indicated with the aid of their respective names, each serum-binding protein preferentially binds a distinctive class of steroid hormones.
Larger dynamic roles beyond steroid hormone delivery. SHBG plays a role in smartphone membrane-related signal transduction through the second messenger cyclic adenosine monophosphate (Camp) and protein kinase A (PKA). Further, cellular-floor SHBG receptors have been recognized. tissues including the breast, testis, and prostate, in addition to assisting a function for SHBG in cellular telephone signaling.
Mechanisms of Steroid Hormone Action
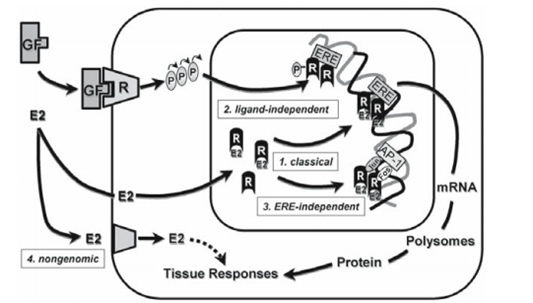
The results of steroids are typically gradual approximately the quick time courses for the effects of 2d-messenger-mediated peptide hormones. that is owing every to the. signal amplification inherent to 2d-messenger cascades and the slower adjustments in gene transcription and translation exerted thru steroids (genomic effects). Early. experiments established those paths of nuclear hormone movement through the usage of protein and RNA synthesis inhibitors which includes cycloheximide and actinomycin D, respectively. Even though the maximum characterized effects of nuclear hormones are mediated with the aid of nuclear receptors and genomic pathways, there are examples of very fast, “non-genomic” outcomes of steroids that display up to be thanks to membrane-mediated outcomes. similarly, the preference mechanisms of nuclear hormone receptor (NHR) activation encompass ligand-independent activation and genomic activation independent of a hormone-responsive detail. so that trans activation domain names of the NHR interact with proteins comprising the transcriptional complicated certain to a promoter and, consequently, stimulate or inhibit rates of transcription.
HREs are families of notably associated DNA palindromic repeats. The estrogen, COUP thing, thyroid hormone, and retinoic acid receptors percentage incredibly homologous consensus response factors, and GR, AR, PR, and mineralocorticoid receptors (MR) percentages very similar, and in some cases, equal factors. The high degree of homology between and within these HREs is likewise contemplated within the high degree of homology between protein sequences of the DNA binding domains (DBD) of the numerous receptors. This would appear to create a hassle with the specificity of hormone action, however, as visible in Table 1, the mutation of nucleotides is enough to regulate a consensus estrogen reaction element (ERE) into a consensus androgen response detail. further, as other non-consensus elements are characterized by using milder being shed on the nature of NHR-precise interactions with the genome a sequence of some characterized response elements for ERs vs ARs, PRs, and corticoid receptors are given. Also provided are consensus sequences for an ERE and a GRE (GRE consensus sequence is identical to a PRE and an ARE). Italicized nucleotides demonstrate potential sites for mutation that can convert one class of 4 to another.

The special lessons of steroid hormones are all present in the movement, and their respective levels range with the one-of-a-kind physiological states of the organism. In addition, there is explicitly more than one target cell.
training of NHR. This provides the organism with them. The problem of the way to spark off a specific gene by using a selected steroid hormone. The specificity of steroid hormone–activated gene expression lies in (1) hormone-precise binding using the receptor, and (2) DNA-unique binding. through the special styles of steroid receptors, and (three) manipulation of get admission of steroid receptors to genes through the differential company of chromatin inside the many goal cells and tissues. among the hormone-insensitivity, syndromes stem from mutations that regulate steroid- or. NHR DNA-binding traits Overall,
NHR proteins are a surprisingly conserved organization of “ligand-structured” nuclear transcription elements (Figure As a whole, NHR proteins are a surprisingly conserved organization of “ligand-structured” nuclear transcription elements (Figure 2). NHRs are modular and can be. broken down into unique practical domain names, which include trans-activating domains, DBD, and LBD. the. extraordinary lessons of NHRs—AR, PR, ER, GR, and MR, the DBD is the most surprisingly conserved location followed by the LBD and then the amino-terminal trans-activating area. the following dialogue of different practical domain names makes a specialty of the ER, but among the traits keep for other NHR kinds.
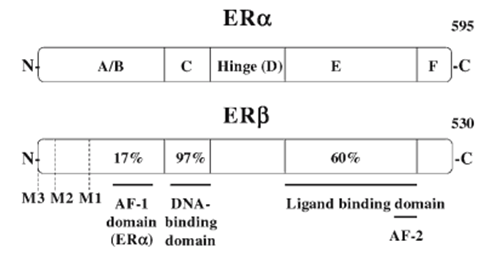
Structure of ER Gene and Protein
ER styles have been identified, ER-α and ER-β, which can be coded via separate genes positioned on separate chromosomes. Each ER protein contains a modular practical domain structure of the steroid hormone nuclear receptor superfamily. The ER proteins contain six purposeful domain names which can be termed A/B, C, D, E, and F domain names. those domains had been observed to possess the following capabilities: ligand-impartial activation characteristic (A/B), DNA binding (C), ligand binding (E), nuclear localization (D), and dimerization and ligand-dependent activation feature E (Fig. 3). The ER-α and ER-β protein percentage is of an excessive degree of homology within their DBDs and LBDs, 97 and 60%, respectively, which ends up in both receptors binding to the same EREs and showing a similar binding affinity for most endogenous and exogenous ER ligands. Then. modular nature of the unique practical domain names and the interdependence of this domain names approach that splice variations of NHR m RNAs can produce altered proteins that behave in appreciably distinctive models from the complete-length NHR. The importance of those versions in regular physiology is still below investigation, however, splice versions can also play a role in ailment states, together with the progression from steroid-established to -impartial most cancers
Ligand Binding domain
ligand-independently and impartial” ER, it is viable that the LBD functions as a repressor of a transcription component that might usually be constitutively energetic. indeed, a constitutively. energetic exon five splice version of ERα has been detected in some human breast cancers. in the end, it seems that E2. binding to the LBD of the ER is not continually vital for ER-mediated genomic actions. recent proof has. proven ligand-independent ER activation of target genes thanks to increasing element activation of kinase signaling pathways.
DNA-Binding Domain
The DBD well-known shows precise binding for sequences of DNA termed EREs. Surprisingly, this region was conserved. Incorporates “zinc finger” motifs, every of which contains cysteine residues that bind zinc. the first zinc finger dictates collection-precise interactions with DNA, and the second one seems to dictate the spacing necessities among the hands of the palindrome. these arms are critical for DNA binding; however, the surrounding amino acids additionally influence binding. The canonical. Element is a palindrome inverted repeat (GGTCAnnnTGACC), even though deviations from this consensus sequence are common (see desk 1). The ER binds. to the DNA collection as a dimer with one receptor molecule contacting each 5-bp inverted repeat. The binding. of the ER-ligand complex to an ERE series roles the ligand-activated ER and associated co activator-activator in which they can interact with basal transcription complexes and affect the cost of gene transcription. In, addition to ERE-mediated gene expression, recent proof shows that the ERs are capable of transacting voting genes whose promoters lack a purposeful ERE via protein-protein interactions with different DNA-bound transcription factors, including Fos and Jun, at AP-1-binding websites. The result of the ER affiliation is. tethering of the ER to DNA and upregulation of gene expression thru an ERE-independent mechanism SRC-1, GRIP1, AIB1, p68 helicase, and CBP/p300. The p160 family of co-regulators contains characteristic α-helical LXXLL motifs that are involved in AF recognition and binding.
Transcription Activation Function
ERα incorporates two regions known to possess transcriptional activation capabilities, activation feature-1(AF-1) and AF-2, placed inside the A/B and E domains, respectively. depending on the cell's kind and goal. genes, AF-1 and AF-2 can act independently or in live performance. As an instance, the removal of AF-1 does now not affect E2. induction of a reporter assembles containing the vitellogenin ERE, while the equal AF-1 poor ERαhas the best 20% of the wild-kind induction of a PS2-ERE.As referred to earlier, the removal of the LBD containing AF-22) can cause a constitutively energetic ERα. this constitutive pastime can also require phosphorylation and activation by way of 2nd messengers. Studies on the use of AF-1 and AF-2 truncated ERα have verified that AF-1 responds to boom elements that act via second messengers, including cAMP or mitogen-activated protein kinase (MAPK) signaling pathway activation, whereas AF-2 is E2 (ligand) dependent. As a result, ER is a nuclear transcription factor that responds to both steroid and 2nd-messenger signaling pathways. In comparison to the nicely characterized activation domains of ERα, the homologous regions of ER-β have not been described in detail regarding transcriptional pastime. “Ligand-impartial” or second messenger activation of transcriptional interest has also been demonstrated for AR and PR, suggesting that this may be an essential and conserved mechanism for the physiologic activation of steroid receptors. The transcriptional activation features of AF-1 and AF-2 a mediated through transcriptional co-regulators, proteins that provide the link among ligand-activated, DNA-certain receptors, and the general transcriptional equipment. The confrontational change brought on through agonist binding to the ER permits co-regulators to have interaction often with AF-2 websites on the receptor; however, interplay with AF-1 websites does occur. Many. co-regulators had been recognized that engage with the ligand-certain ER, which includes the p160 family members SRC-1, GRIP1, AIB1, p68 helicase, and CBP/p300. The p160 circle of relatives of co-regulators carries featureα-helical LXXLL motifs, which can be involved in AF recognition and binding.
Dimerization
Most facts suggest that NHRs act as homodimers, although a few data suggest viable consequences of NHR monomers. The vicinity of the protein was answerable. dimerization of the mouse ER overlaps with steroid binding characteristics and spans amino acids 501–522. these. amino acids shape an amphipathic, helical structure with a less-than-perfect heptad repeat of hydrophobic amino acids reminiscent of the leucine zippers found in the JUN/ FOS and CREB households of transcription factors. Mutations of amino acids on this hydrophobic stretch have verified that this area is vital for dimerization, steroid binding, and, as a result, trans activation. The dimerization. The tend characteristic is vital for the results of NHR homodimers but may also play a role in the formation of heterodimers between NHRs and other transcription elements. Heteroatoms including ERα and ER-β, in addition to heterodimers of SP1 proteins, had been shown to modify the expression of genes inclusive of c-FOS and trans-forming increase component-α. hence, the dimerization function is important for the results of NHR homodimers however also performs a role in the formation of heterodimers between NHRs and other transcription factors with comparable dimerization domain
Nuclear Localization Signal
NHRs and plenty of various transcription factors private a segment of amino acids that goals the proteins within the cell nucleus. those stretches of amino acids have a tendency to be primary and have been termed the nuclear localization sign (NLS). The NLS is located between them. amino acids 250 and 270 of the ER? a region that stocks homology with the nuclear localization domain names of the glucocorticoid and progesterone receptor expression, respectively. The NLS for. ER? has but to be characterized.
Nongenomic Mechanisms of Steroid Movement
Despite the truth that steroids usually act through the traditional genomic mechanism, a method that takes numerous mins to hours for outcomes to be seen. Steroids also can elicit rapid biologic effects within seconds to minutes after control through nongenomic mechanisms. Nongenomic steroid movement outcomes. inside the rapid activation of a spread of cell-signaling molecules, such as MAPKs, adenylyl cyclase; PKA, and %. Fast responses to estrogen have been previously reported. determined in granulosa cells, endometrial cells, and oocytes, all of which show off advanced intracellular calcium concentrations hastily, if no longer without delay, After E2 exposure. different estrogen-mediated non-genomic. mechanisms had been found in spermatozoa, breast cells, nerve cells, and vascular tissues. similarly, non-genomic mechanisms had been defined for progesterone, androgens glucocorticoids, and mineralocorticoid. current research is underway to decide whether or not those non-genomic steroid mechanisms. our thanks to receptor impartial activities at the plasma membrane, nonsteroid-associated membrane receptors, or membrane-certain NHRs.
Steroids and Development
Scientists have seemed for years that during utero and neonatal publicity steroids are critical for sexual differentiation of the brain and peripheral reproductive structures. A guiding idea for the examination of increasing intellectual moves of steroidal consequences is the company activation speculation. stated definitely, prenatal or neonatal exposure to steroid hormones organizes or alters differentiation of the phenotype such that hormonal exposure in adulthood is more likely to activate a specific reaction. A corollary of this rule is that the preliminary exposures must fall into interior, high-quality critical periods of sensitivity. those essential durations commonly occur at some unspecified time in the future of Fetal, neonatal, and pubertal degrees. Steroids affect the improvement of organs and tissue via induction and inhibition of the boom. Inhibition takes vicinity through active cellular demise, a technique termed. apoptosis. Apoptosis is a lively technique requiring protein synthesis and ensuing in chromatin condensation, degradation of chromatin in a function-segmented manner that produces an observable “ladder” pattern and development of apoptotic bodies.
Stromal-Mesenchymal Interactions
A recurring theme in the development of steroid-dependent glandular tissues is the Importance of stromal mesenchymal tissue induction. In this scheme, fate is. of undifferentiated epithelium is determined by the underlying mesenchyme with, which comes into contact. For instance, undifferentiated epithelium combinedwith prostatic or integumental mesenchyme. develops a phenotype dictated by the type of men chyme. In the case of hormone-directed-indicates morphogens such as in the prostate or breast, hormonal. influences on the glandular epithelium can occur Either directly on epithelial cells or indirectly via inductive influences of the mesenchyme. Recent experiments. demonstrate that epithelium can also influence the underlying mesenchyme, indicating a bidirectional epithelial-mesenchymal interaction.
Secondary Sex Structures
In the developing mammalian embryo, gonadal sex is determined based on the useful resource of the genotype. In flip, the e Behavior embryonic. gonads secrete hormones that, coupled with maternal hormones, decide the early hormonal milieu to which secondary sex systems are exposed and, consequently, dictate the development of male or female phenotype. Dogma holds that mammals have a default device such that embryos grow a girl's phenotype in the absence of any gonadal steroid hormones. In guys, as the developing testes begin to increase sex cords and secrete Müllerian-inhibiting materials (MIS) and testosterone. The MIS induces ipsilateral regression of the Müllerian ducts, which prevents the improvement of Müllerian derivatives together with the uterus and Fallopian tubes. prolonged testosterone stimulates the improvement of Wolffian derivatives which include the epididymis, vas deferens, and seminal vesicles. Differentiation of outside genitalia and accessory glands (inclusive of the prostate) from. The genital tubercle, scrotal folds, and urogenital sinus
Sexual behavior and Sexual Dimorphisms of the Brain
intercourse conduct in most adult vertebrates is dependent on (1) organizational results of hormones early in improvement, and (2) activation consequences of circulating steroids within the person. in lots of species, in-utero and neonatal hormone exposures alter adult patterns of sexual behaviors. traditionally, this remark caused the belief that to some organizational degree, the brains of men and girls must be morphologically or functionally awesome to desire girl- or male-typical behaviors. In the case of the rat, sexually dimorphic nuclei have been found inside the critical fearful gadget (CNS). Male rats possess enlarged sexually dimorphic nuclei inside the medial preoptic region of the hypothalamus and within the spinal cord. The development of these nuclei and subsequent characteristics in adult guys is androgen-dependent; androgen ablation during early crucial periods of differentiation leads to smaller, lady-typical nuclei and decreases in male-commonplace copulatory conduct. In rats, testicular testosterone at the sexually dimorphic nuclei of the medial preoptic location appears predominantly through aromatization to E2; treatment with E2 mimics the impact of testosterone, and the use of an aromatase inhibitor can prevent the masculinization of sexually dimorphic nuclei. Comparable steroid-dependent Dimorphisms have been observed inside the CNSs of gerbils, voles, songbirds, lizards, and fish. Dimorphisms may be gifted as versions of gross quantity, cellular variety, cellular size, dendritic arborization, and ranges of expression of enzymes, neurotransmitters, Neuropeptides, or receptors. Sexual disorders have also been reported in humans. reported within the anterior hypothalamus (AH), preoptic area (POA), and anterior commissure, although there are some conflicting facts.
Steroids and Bone
Bone cells express ER, AR, and PR, and the development and maintenance of bone structures are regulated by estrogen and androgen. Pubertal surges in estrogens and androgens initiate growth spurts including long bone growth, primarily mediated by increased insulin-like growth factor-1, and, subsequently, cessation of bone Growth through epiphyseal closure. In adults, E2 maintains bone mass and mineralization. The importance of the effects of E2 on bone growth and development is evident in individuals lacking E2. For instance, A human male patient lacking functional ERα exhibited continued bone growth, decreased bone density, and absence of epiphyseal closure (see Section 6.3). In addition, the absence of E2 due to ovariectomy or menopause contributes to osteoporosis, whereas exogenous E2 ameliorates this condition. Excess production of cortisol results in a loss of bone mass (osteopenia)
Steroids and Liver
Liver cells specific ERs and ARs regulate the manufacturing of secreted proteins and steroid-metabolizing enzymes15-fold Lowe. In humans, the liver synthesizes and secretes the liver. into the bloodstream a plasma protein termed SHBG. This protein serves to sequester and save your steroids from being metabolized and/or cleared from the bloodstream. SHBG binds DHT with excessive affinity (kd ~ 0.5nM) and testosterone and E2 with approx 5- and 15-foldlower affinity, respectively. Estrogens stimulate.
Androgens inhibit the synthesis and secretion of hepatic SHBG
There are awesome intercourse variations inside the profile of Steroid metabolites are excreted in the urine. The basis of such sex variation is the consequence of sex differences in the expression of metabolic enzymes in the liver. As an example, the lady's liver expresses the 15α-hydroxylase activity.
However, this enzyme is not specific to males. In contrast, adult males have explicit sixteen-hydroxylase, which is absent in females. In rats, those sex variations are regulated by what constitutes a hypothalamic-pituitary-hepatic axis, wherein neonatal androgens masculinize the boom hormone (GH) axis. In flip, the sample of (GH) secretion imprints a male or girl profile of steroid metabolism. that is glaring through the reality that pulsatile surges of GH (male-like sample) or tonic, low-stage GH infusions (female-like) into hypophysectomies rats produced a male- or female-typical pattern of enzyme expression and metabolism, respectively. STEROIDS AND Normal physiology
HPG Axes
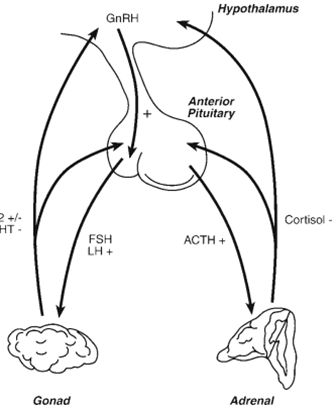
The gonadal function is regulated by pituitary gonadotropins (LH and FSH), which may be regulated by the hypothalamic peptide, gonadotropin-releasing hormone (GnRH), steroids, and gonadal peptides (Fig. four). GnRH is synthesized by small populations of neurons within the POA, AH, and medio basal hypothalamus, and is released into the hypophysial portal machine, which incorporates GnRH into the pituitary. GnRH stimulates synthesis and launch. Of LH and FSH, which, in turn, modify steroidogenesis and gametogenesis. Elevations within the circulating concentrations of gonadal steroids provide feedback on the hypothalamic and pituitary websites to adjust gonadotropin levels. Synthesis and release. In addition to steroids, gonads secrete peptide hormones (activin and inhibin) that feed again in the pituitary to modify the synthesis and launch of gonadotropin.
GnRH secretion takes place as both an episodic and tonic pattern and as a surge associated with ovulatory events in females. The tonic pattern of GnRH secretion happens in pulsatile style with a periodicity of approximately 1 pulse/h. Steroids feed lower back on the GnRH pulse generator concept to be located in the medial basal hypothalamus to regulate tonic patterns of GnRH secretion. law of the ovulatory GnRH surge, however, appears to require input from the POA/AH areas. further, androgens and estrogens can feed lower back directly on pituitary gonadotropin to alter cell boom, sensitivity to GnRH, and basal stages of gene expression of gonadotropins. A final level at which steroids feedback is on steroidogenic cells themselves. for example, experiments imply that androgens can down-regulate LH-precipitated expression of steroids by using Leydig cells. The. comments effects of steroids on these extraordinary levels constitute a lengthy loop (gonad-hypothalamic), brief loop(gonad-pituitary), and ultrashort loop (gonad-gonad) comment circuit (Figure 4).
Female Reproductive Cycles
Human beings and the maximum different girl mammals are spontaneous ovulatory, that is, the cyclical buildup of estrogen triggers a “spontaneous” pulse of gonadotropin that triggers ovulation unbiased of mating stimuli. however, in a few species, females are reflex ovulatory; that is physical mating stimuli are chargeable for triggering GnRH and gonadotropin surges provide the proximate cues for ovulation. the subsequent discusses normal ovarian cycles in spontaneous ovulatory. As a follicle matures underneath the impact of basal degrees of FSH, circulating tiers of E2 growth to a topic or near ovulation. The increase in E2 will increase. gonadotropin sensitivity to GnRH with the aid of up-regulating producers GnRH receptors and, at height levels, experts' nice remarks impact that triggers a GnRH surge that, in flip, produces an LH surge. This LH surge induces ovulation, the formation of corpora lutea from granulosa cells of the follicle, and the synthesis and secretion of progesterone. E2. Over the route of the ovarian cycle, ovarian steroids exert control over GnRH and gonadal tropine, maturation of follicles, and practice of the uterus for Implantation, changes in vaginal and cervical function, and conduct remarks. A lengthy ovariectomy of mice leads to a large growth in consistent-nation messages for FSH and LH, whereas estrogen remedies the opposite effect through a terrible remark mechanism. However, this is an oversimplification of the complex. Outcomes of ovarian steroids on the feedback law of the hypothalamus and pituitary. Estrogen seems to have. biphasic consequences on the synthesis and secretion of LH and FSH in which decreased degrees of E2 is present throughout post-ovulatory and early follicular levels can suppress gonadotropins, while the better ranges of E2 discovered throughout overdue follicular genesis bring about (1) extended sensitivity of gonadotropin to GnRH, and/or (2) a pre ovulatory pulse of GnRH. in addition, following E2 priming, the initial exposure to progesterone results in improved sensitivity to GnRH accompanied with the aid of lengthy-time period inhibition. certainly, extended progesterone related to the formation of the corpora lutea results in the suppression of estrous cycles.
Effects of Estradiol and Progesterone on Accessory sex structures. As E2 ranges increase throughout. the follicular phase, the luminal epithelium of the uterus enters a proliferative segment in training for implantation. In mice, initial E2 publicity swiftly induces hyperemia and water inhibition (1–four h), followed by a boom in DNA and protein synthesis, hyperplasia, and hypertrophy. The crucial impact of E2 at this level is. Induction of progesterone receptor synthesis, which permits the uterus to reply to multiplied progesterone.
in the course of luteal stages, the uterus enters a proliferative section throughout which improved progesterone completes the coaching of the endometrium for implantation of the blastocyst through growing vascularization and with the resource of thickening the mucosal layer of the epithelium. inside the absence of implantation, the corpora lutea degenerate, serum progesterone degrees drop, and the endometrium degenerates. The hypothalamic-pituitary-gonadal axis is then freed from progesterone suppression to renew every spherical folliculogenesis.
Mammary gland characteristic is regulated with the resource of the coordinated moves of estradiol, progesterone, and prolactin (PRL). Estradiol promotes lobuloalveolar improvement by appearing immediately away from the mammary gland and by stimulating the synthesis and secretion of PRL through the use of the anterior pituitary. Estradiol-stimulated increase in PRL
assisted in the preparation of the glandular tissue for lactation. Progesterone promotes glandular development but requires (1) the presence of pituitary hormones and (2) priming with E2, which regulates-regulates stages of PR. whilst p progesterone. and E2 help put together the glandular tissue for lactation, those hormones additionally suppress lactation until Parturition expulsion of the placenta causes an abrupt decrease in E2 and progesterone Puberty. crucial ranges of intercourse willpower and sexual differentiation arise in utero and early neonatal life. However, terminal differentiation of sexual best-friend dimorphic systems and the onset of reproductive fertility occur during puberty. In humans, the onset of puberty is marked by a boom in tonic, pulsatile GnRH launches, and extends the secretion of LH and FSH.
Inside the lady, extended gonadotropin levels provoke waves of folliculogenesis and a related increase in E2 and androgen levels. As levels of E2 boom, terminal varies initiation of the breasts starts off evolved and females undergo a boom spurt. As E2 tiers boom over the route of puberty, E2 induces epiphyseal closure and cessation of the boom spurt. exposure to increasing stages of E2 results in an initial proliferation of the endometrium, followed by primary menses (menarche) as a result of a drop in E2 on the abandonment of a follicular wave. The preliminary ovulatory occasion takes approximately one-year following menarche, presumably because the mechanisms regulating a GnRH surge now reply to E2-superb remarks. inside the male the pubertal onset of increased GnRH and gonadotropin synthesis and launch is marked by testicular amplification and initiation of spermatogenesis and steroidogenesis. As ranges of circulating testosterone growth, penile enlargement, the boom of pubic hair, and boom spurts start. further, the glandular epithelium of secretory glands which include the seminal vesicle and prostate undergo a proliferative segment and begin to supply secretory merchandise that grows to be additives of the semen.
Results of E2 and Progesterone on Sexual Behaviour
In many vertebrates, E2 and progesterone act to coordinate periods of maximum sexual receptivity with periods of the most likelihood of fertilization. As a result, E2 priming throughout follicular stages followed via a surge of progesterone associated with ovulation and luteinization consequences in most receptivity near the time of ovulation. The consequences of the estrous cycle on conduct may be re-created in ovariectomized lady rodents treated with E2 followed by progesterone treatment. The E2 remedy has a facilitatory effect alone but is substantially augmented through the next progesterone remedy. primarily based on lesion research and intra-hypothalamic implants of E2 and progesterone, the ventromedial hypothalamus seems to be the web page of E2 and progesterone outcomes on Receptive and perceptual behaviors in female mammals. One impact of E2 is to upregulates PR in the ventromedial hypothalamus. The significance of E2 and progesterone in the sexual behaviors of female humans appears to be much less seasonally determined than that in other mammals with awesome behavioral estrus.
Male Reproductive Cycles
requirement of the male HPG pivot is five public tons less vital a method in which GnRH pulses are dropped in account and do not endure surges like the one guiding ovulation in wives. In rodents, the male GnRH device cannot put oneself in the place of another external E2 accompanying a surge; concurrently with activity in persons and monkeys, injections of exogenous E2 cause success in a GnRH surge. On account of this, the omission of a GnRH surge in males is due to the lack of estrogen development to guide folliculogenesis in adolescents.
GnRH induces gonadotropins to combine and release FSH and LH, which individually participate in spermatogenesis and steroidogenesis. LH elevates cAMP levels, which provokes the combining and discharge of test oestrone from the Leydig containers. extended testosterone assists spermatogenesis and feeds again to downregulate GnRH stages and, because of this, combination and release of Gona drop-in. testosterone can furthermore additionally feed repeated all right away as an androgen and as an estrogen remark insult aromatization. certainly, present studies accompanying αERKO adult men attach that ERα further to the AR road can favorably suppress the antitoxin LH at the pituitary and hypothalamus levels. Gonadotropins particularly each AR and ERα event GnRH neurons do inexact mammoth portions of AR or ERα. For that reason, the hypothalamic response occurs indirectly through AR- and ERα-signifying neurons that stimulate the interest of GnRH neurons from a much-off net location on the world wide web. Sexual performance in maximum-type male vertebrates is contingent on the comprehensive flow of testosterone tiers. Testosterone acts at the idea’s nuclei of the AH/POA to prompt male-common intercourse actions; lesions of those mind regions or emasculation effects in the ending of intercourse behaviors. relying on the class, observable consequences of androgens grant permission to be a habit to all aromatization accompanying E2 and direct outcomes in addition to testosterone or DHT. Testosterone acts more on Sertoli containers, which concedes the possibility of continued spermatogenesis even in hypophysectomies fellows. any unique detracting feature is the stimulation of ornamental sexuality plans that amount to the prostate and generative sac. Circulating testosterone is molded into fiveα-DHT, which causes hypertrophy of the secretory epithelium, progresses RNA and protein combining, and leads to superior protein secretion. As persons, the persistent accompanying functioning of androgen-generally located answers is located entirely on uncovering flowing testosterone.
HPA Axis
The synthesis and secretion of glucocorticoids such as cortisol are stimulated by the pituitary hormone ACTH. The release of ACTH from corticotropes is stimulated by a hypothalamic peptide, corticotropin-releasing factor (CRF), produced by the paraventricular nucleus of the hypothalamus. Increased levels of cortisol act directly at the level of the corticotrophin to reduce ACTH production and at the para-ventricular nucleus of the hypothalamus to suppress CRF levels (Figure 4). Thus, hypophysectomy leads to a decrease in cortisol levels, and adrenalectomy leads to an increase in ACTH levels, which is reversed by cortisol treatment. Continuous. ACTH secretion, characteristic of chronic stress, also leads to adrenal cortical hypertrophy, elevated glucocorticoid levels, and, in some cases, adrenal failure. ACTH has a very limited tropic effect on the zona Glomerulosa and aldosterone levels in mammals. Primary regulators of aldosterone synthesis include potassium (K+), sodium (Na+), blood volume, and angiotensin II. Elevated angiotensin II and K+ loading and low serum Na+ levels increase the synthesis of adrenal aldosterone, which promotes Na+ reabsorption by the kidney. Conversely, Na+ loading causes hypertension or circulatory expansion and decreases aldosterone levels. Potassium appears to act directly at the level of the glomerulosa both in vivo and in vitro. Furthermore, K+ loading appears to affect the release of renin and synergizes with angiotensin II to increase the release of aldosterone. Sodium depletion decreases the blood volume, which stimulates the juxtaglomerular apparatus to secrete renin. In turn, renin cleaves Angiotensinogen, producing angiotensin I, and initiating a cascade. eventually leads to elevated angiotensin II levels and subsequent elevation of aldosterone levels. Elevated aldosterone levels promote Na retention and elevate blood levels. volume and arterial pressure, which, in turn, feeds back to decrease renin production.
Carbohydrate Metabolism
Adrenalectomy leads to reduced liver glycogen and low blood glucose levels, resulting from increased oxidation of glucose and decreased gluconeogenesis from proteins.
Conversely, the administration of cortisol leads to a rise in blood sugar and an increase in liver glycogen stores owing to decreased glucose utilization and increased gluconeogenesis. Glucagon also elevated glucose levels. and promote glycogen breakdown. Insulin, however, has the opposite effects, producing lower blood sugar (decreased gluconeogenesis), and increased glycogen synthesis and storage.
Stress Responses
Stress can be induced through social interactions, physical stress, and physiological challenges. Classic stress. The response is the increased secretion of glucocorticoids triggered by the increased secretion of CRH and ACTH. Hormonal responses to stress can be very fast (minutes) and dissipate quickly, or in some cases, become chronic. In stressful situations, elevated glucocorticoid levels stimulate an adaptive increase in glucose levels. for carbohydrate energy sources. The effects of long-term elevated glucocorticoid include suppression of HPG function and suppression of the immune System Elevated glucocorticoids suppress the production of GnRH by hypothalamic neurons and, consequently, alter gonadotropin and steroid synthesis, and gametogenesis. Glucocorticoids also directly affect gonadotropin function by suppressing basal and second messenger-induced synthesis and release of gonadotropins. At the level of the gonads and glucocorticoids. also suppresses gonadotropin and second messenger-stimulated steroid synthesis. Thus, the suppressive. The effects of elevated glucocorticoids on reproduction are exerted at all levels of the HPG axis.
Glucocorticoids play a role in apoptotic events leading to the differentiation of the immune system and regulation of the immune system in adults. Apoptosis is an active process of programmed cell death, in which a series of programmed events leads to cell death. Glucocorticoids are used for the development of the immune system. induces apoptosis in auto-reactive T-cells and un reacted B-cells. High levels of glucocorticoids can also lead to apoptosis of immune cells in the adult and, consequently, a compromised immune system and increased susceptibility to disease
Electrolyte Balance
Salt balance is achieved primarily by mineralocorticoid and neurohypophysis peptides such as arginine vasopressin and arginine vagotonic. Aldosterone, the Principal human mineralocorticoid reduces Na+ loss by enhancing resorption in the renal tubules of the kidney. Thus, adrenalectomy or deficiencies in adrenal steroid synthesis result in rapid decreases in blood Na+ levels and circulatory collapse unless Na+ or exogenous aldosterone is provided.
Steroids and Pathophysiology
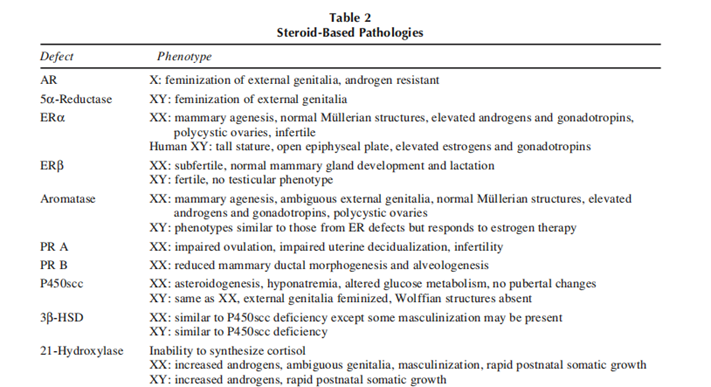
Steroid-associated pathologies (table 2) encompass non heritable steroid-structured cancers and heritable syndromes that affect the synthesis or feature of steroids and their receptors, resulting in steroid hypersensitivity syndrome. Although the outcomes of steroids in steroid-established cancers are environmental and non-heritable, there are genetic predispositions to them. growing such cancers. Heritable defects in steroid motion are normally autosomal recessive syndromes that cause developmental anomalies of various tiers of severity.
Cancer
Several steroid-dependent and steroid-independent tumors occur in steroid-target tissues such as the uterus, breast, and prostate. In the case of prostate cancer, a clear link with androgens is provided by the fact that castrated males never develop prostate cancer. Furthermore, many prostate cancers exhibit a period of regression and remission following castration and anti-androgen treatment. Unfortunately, many of these cancers enter a steroid-independent stage during which growth and metastases are independent of androgens or hormonal therapy. A vital question is, why does prostate cancer become steroid independent? Since normal proliferation and growth cycles are dependent on androgens, the question becomes, why do these tumor cells lose their normal requirement for androgen stimulation? Two hypotheses seem viable: (1) splice variants result in a constitutively active variant AR that stimulates growth independent of androgens, and (2) key regulatory points in the cell cycle lose the requirement for androgen stimulation. These hypotheses remain to be tested in this and other steroid-independent tumors.
Breast cancer is often amenable to treatment with tissue-specific steroid antagonists, including tamoxifen and faslodex, as assessed clinically by assays for both ER and PR in mammary biopsies. The presence of receptor levels >10–15 fmol suggests that the cancer is probably steroid-dependent and likely to respond to antihormone therapy. However, breast cancer can become estrogen-independent and unresponsive to antiestrogens such as tamoxifen. A constitutively active ERα splice variant present in some breast tumors may provide one explanation of how cancers can progress to a steroid-independent state. Another explanation may be that the overexpression of ER co-activators, such as amplified in breast cancer-1, in breast cancer cells result in increased levels of co-activator activity that could reduce the effectiveness of ER antagonists such as tamoxifen.
Androgen-Based Developmental Defects
Several types of androgen-based defects have been documented. These range from defective. The 5α-reductase enzyme occurs as a rare autosomal mutation to a defective AR resulting from mutations within the X-linked AR gene. In addition, alterations in. steroidogenic enzymes earlier in the synthetic pathways can also result in developmental anomalies of androgen-dependent tissues. The phenotypic manifestations. of these defects range from infertility in phenotypically normal males to complete feminization of external genitalia. In cases of enzymatic deficiencies, hormones. therapy can ameliorate some symptoms, whereas those symptoms related to receptor defects are resistant to hormone therapy.
5α-Reductase Deficiency
5α-Reductase type 1 is expressed at low levels in peripheral tissues, and 5α-reductase type 2 is expressed at high levels in male genital structures. In males, 5α- reductase deficiencies result in varying degrees of ambiguity of the external genitalia ranging from hypospadias to complete feminization. Under the influence of elevated testosterone, Wolffian derivatives such. as the epididymis and seminal vesicle develop normally whereas the external genitalia are feminized to varying degrees. In addition, Müllerian derivatives are absent. owing to the production of MIS by the testes. In extreme. cases of feminization, this syndrome is often diagnosed at the age of puberty when a patient with female phenotype exhibits amenorrhea and/or some increased mas colonization owing to the increased levels of testosterone associated with puberty. Before puberty, these individuals are usually raised in female gender roles, but. following pubertal changes in phenotype, they sometimes assume male gender roles.
Androgen Insensitivity Syndrome and Testicular Feminized Males
Androgen insensitivity presents itself as a spectrum of disorders, ranging from complete external feminization to infertility in phenotypic males. A wide. variety of AR gene defects have been documented, ranging from point mutations that cause a premature stop codon in the testicular feminized male mouse to a complete deletion of the AR gene in a human family. Known. mutations within the human AR appear to cluster primarily within the DBD and SBD of the receptor. Generally, there is a reasonable correlation between the degree of feminization and the degree to which the normal function of the AR is altered, as assessed by various in vitro assays. For instance, mutations that abolish steroid binding led to profound feminization, and more subtle mutations affecting thermolability and steroid dissociation rates lead to less profound effects, such as infertility and hypospadias. Fertility problems related to AR defects are resistant to therapy, whereas anomalies such as mild hypospadias can be treated by surgical correction. In cases of. complete feminization, inguinal and labial testes are removed owing to increased incidences of testicular cancer. Infertile completely feminized XY individuals. develop female gender roles and tend to maintain these roles throughout adulthood.
Estrogen-Based Developmental Defects
Until recently, no mutations in the aromatase or ER genes had been detected. Additionally, ERα mRNA had. been detected during very early embryonic stages using reverse transcriptase-polymerase chain reaction. Thus, it was suspected that estrogen is critical for the development of a viable embryo and that mutations of either of the aforementioned genes would be lethal. Recent findings, however, have documented aromatase deficiency. and estrogen insensitivity (ERα defects) in adult humans. In addition, gene-targeted mouse lines in. which ERα(αERKO), ERβ (βERKO), or both ERα and ERβ have been disrupted (αβERKO) demonstrate that embryos can develop in the absence of functional nuclear ER. Furthermore, aromatase-deficient mice. (ArKO), which lack the enzyme for converting androgens to estrogens, and therefore have no circulating estrogens are viable. Although these data suggest that estrogens may not be critical for embryonic survival, several phenotypic and receptor-specific abnormalities occur owing to these gene mutations.
Aromatase Deficiency
Mutations in the aromatase enzyme led to alterations in phenotypes in both males and females. A male homozygous for defective aromatase exhibited tall stature, incomplete epiphyseal closure, continued linear bone growth, and osteoporosis. Circulating androgens and. gonadotropin levels were increased, but gross sexual phenotype was normal. In an aromatase-deficient female, the individual presented with ambiguous genitalia at birth but normal internal Müllerian structures by subsequent laparoscopic examination. At puberty, the individual possessed the following symptoms: absence of. breast development (mammary agenesis), primary amenorrhea, elevated gonadotropins, elevated androgens, and polycystic ovaries. Estrogen treatment alleviated many of these symptoms. The masculinization is owing to a lack of conversion of C19 steroids into estrogens and, hence, excess circulating androgens. As discussed previously, ArKO mice have been generated to characterize further the phenotypes associated with aromatase and estrogen deficiency. Many of the phenotypes observed in ArKO mice are similar to those seen in aromatase-deficient humans. Male ArKO. mice exhibit an osteoporotic phenotype owing to decreased bone formation, increased adipose tissue, elevated gonadotropin levels, age-dependent disruptions of spermatogenesis and fertility, and impaired sexual behavior. Female ArKO mice, like males, exhibit an osteoporotic phenotype and increased adiposity, as well as increased gonadotropin level Reproductive phenotypes in the female ArKO mice include underdeveloped external genitalia, uteri, and mammary glands, and infertility.
Estrogen Receptor Mutations
Recent work documented a normally masculinized, human male with clinical symptoms very similar to those of the aromatase-deficient male: tall stature, incomplete epiphyseal closure, osteoporosis, decreased sperm viability, and elevated testosterone and gonadotropins. Estrogen levels were also elevated and the patient exhibited no response to E2 therapy. Molecular. analysis revealed a point mutation that created a premature stop codon in the ERα gene, resulting in a truncated mutant form of the receptor protein. ERKO mice for both ERα and ERβ have been developed to elucidate the role of ER-mediated signaling in normal growth and development. The female. αERKO mouse shows several interesting phenotypes, including reduced uterine development, absent uterine responsiveness to E2, mammary agenesis, hemorrhagic cystic ovaries, ovulation, elevated gone drop-ins, and elevated testosterone and E2. Male αERKO mice exhibit normal gross phenotype but are infertile as a result of reduced intro missions, reduced sperm counts, and decreased sperm motility. The gross. sexual phenotype of the external genitalia and internal androgen-dependent structures appear normal except testicular dysmorphogenetic results in inning in reduced testis size. In contrast to the complete. infertility in αERKO mice, βERKO females are sub fertile, have normal mammary gland development, and lactate normally, and males are fertile and have no testicular phenotype.
Progesterone-based Developmental Defects
Defects in progesterone synthesis can arise attributable to mutations in P450scc and 3-HSD. but, because. of the pivotal role of progesterone inside the artificial pathways leading to other steroids, the outcomes of the absence of progesterone synthesis are clouded with the aid of the absence of other vital steroids. hence, the effects of defects in progesterone movement may be more. appropriately elucidated from instances related to a faulty PR. To this quit, a PR knockout (PRKO) mouse that was a whole knockout of each isoform of the PR (PR-A and PR-B) was evolved and characterized. the two. isoforms of the PR, PR-A (81 KDA) and PR-B (116 KDA) are expressed from the same gene containing opportunity translation begin sites. girl PRKO mice had been ovulatory, possessed underdeveloped mammary glands, and did not display lordosis conduct. but, estrogen. treatments did motivate uterine expansion, hyperplasia, and edema, indicating a useful ER machine. The male PRKO mouse was fertile and grossly ordinary except for an underdeveloped preputial gland. to elucidate the jobs of the person's PR isoforms, PR-A and PR-B knockout mice, specific PRAKO and PRBKO, respectively, had been developed and characterized. Phenotypic analyses have discovered that: Women PRAKO mice are infertile because of intense abnormalities in ovarian and uterine function. but, PR-A-poor mice have regular mammary. gland and thymic responses to progesterone. with the aid of comparison, PRBKO mice display everyday ovarian, uterine, and thymic responses however exhibit decreased mammary ductal morphogenesis. Taken together, these observations illustrate the awesome roles of each of the PR. isoforms in progesterone motion
Corticoid-primarily based Developmental Defects
Congenital adrenal hyperplasia is a heritable disorder in which the adrenal does not synthesize cortisol efficaciously. the dearth of capacity to synthesize cortisol can the result of defects in any of the enzymes concerned within the synthesis of cortisol from LDL cholesterol. for this reason. of these problems, comments inhibition of the pituitary is absent and excessive ranges of ACTH are secreted, ensuing in hypertrophy of the adrenal and, relying on the affected enzyme, immoderate levels of precursors for cortisol synthesis. A large buildup of cortisol precursors can bring about the synthesis of extra androgens and the next masculinization of females.
A deficiency in LDL cholesterol desmolase (P450scc) affects a deficiency in all steroid hormones and a syndrome known as congenital lipoid adrenal hyperplasia. Deficiency in 3-hydroxysteroid dehydrogenase also affects in an ailment wherein the synthesis of corticoids and sex steroids is terrible within the adrenals and gonads. the one's problems are characterized by an incapacity to produce cortisol and aldosterone and, consequently, a decreased functionality to alter glucose metabolism, an incapacity to hold salt and intense hyponatremia. In men, the absence of intercourse steroids influences feminized outside genitalia, and secretion of MIS with the aid of inguinal testes causing regression of Müllerian derivatives.
ladies with P450scc deficiencies are normal in look at the beginning however do no longer undergo pubertal changes. ladies with 3β-HSD deficiencies may show some masculinization of outside genitalia attributable to the secretion of dehydroepiandrosterone, a susceptible androgen. commonly, the treatment of those issues involves. the alternative of glucose- and mineralocorticoid hormones observed thru sex steroid remedy near pubertal age.
The most common form of adrenal hyperplasia is owing to a deficiency in 21-hydroxylase. The resulting. inability to synthesize cortisol leads to a buildup of precursors and subsequent conversion into testosterone. Clinically, females often present with ambiguous genitalia, irregular menstrual cycles, and some virilization.
All sexes undergo rapid somatic growth (postnatal), accelerated skeletal growth, and early closure of epiphyseal plates. Some patients also have defects in the ability to synthesize aldosterone, resulting in a “salt-wasting” form of the disease.
If the fetus is diagnosed prenatally via genotyping of biopsy samples, the mother can undergo treatment with dexamethasone to suppress the excess production of androgens by the adrenal cortex of the fetus, thus reducing the masculinization of female offspring. Neonatal screening for 21-hydroxylase can be accomplished by assaying for 17-hydroxyprogesterone. This may be useful. preventing deaths related to salt-wasting forms of 21- hydroxylase deficiency.
Methodology:
Describe the research methodology employed, including the study design, participant characteristics (if applicable), and laboratory techniques utilized.
Specify the types of steroid hormones investigated.
Results:
Summarize the key findings of the research, focusing on the specific outcomes related to the research question(s) or hypotheses.
Present any statistical analyses or significant correlations observed.
Discussion:
Interpret the results in the context of existing literature and theories.
Discuss the implications and significance of the findings.
Address any limitations or potential biases of the research.
Consider alternative explanations or factors that may have influenced the results.
Identify potential avenues for future research and areas that require further investigation.
When discussing specific research findings, it is important to consider the diverse functions of steroid hormones in the body. For example, research on cortisol, a glucocorticoid hormone, has examined its role in stress response, immune regulation, and metabolism. Studies on sex steroids, such as estrogen, progesterone, and testosterone, have explored their effects on sexual development, reproduction, and behavior.
It's worth noting that research on steroid hormones is a vast field, covering numerous subtopics and specific hormone functions. To provide a more detailed discussion, it would be helpful to specify a particular aspect or research study related to steroid hormones that you would like to explore further.
Conclusion
Steroid hormones are synthesized and secreted by the ovary, testis, adrenal, and placenta. The majority of site steroidal effects occur through binding to specific intracellular receptor proteins that regulate gene transcription. in target tissues. Steroids are critical for the sexual differentiation of different target organs and, in the adult, are. important regulators of many aspects of normal physiology. Thus, heritable deficiencies in steroid synthesis. or receptor action often leads to permanent alterations in the differentiation of adult phenotype and, hence, altered function in the adult. Steroid-dependent tumors can. become steroid independent by somatic mutations in receptor genes and alternate splicing of m RNAs. Recent advances in molecular endocrinology and gene targeting to generate relevant experimental animal models have allowed scientists to begin to elucidate the molecular mechanisms by which steroid hormones regulate normal physiology and pathophysiology.
Acknowledgment
The completion of this research project would not have been possible without the contributions and support of many individuals and organizations. We are deeply grateful to all those who played a role in the success of this project.
I would like to thank My Mentor [Dr. Naweed Imam Syed Prof Department of Cell Biology at the University of Calgary and Dr Sadaf Ahmed Psychophysiology Lab University of Karachi for their invaluable input and support throughout the research process. Their insights and expertise were instrumental in shaping the direction of this project.
Declaration of Interest
I at this moment declare that I have no pecuniary or other personal interest, direct or indirect, in any matter that raises or may raise a conflict with my duties as a manager of my office Management
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Financial support and sponsorship
No Funding was received to assist with the preparation of this manuscript
References
- Clark JH, Mani SK. (1988). Actions of ovarian steroid hormones. In: Knobil E, Neil J, eds. The Physiology of Reproduction, vol. 1. New York, NY: Raven, 1011–1059.
View at Publisher | View at Google Scholar - Clark JH, Peck EJ, eds. (1979). Female Sex Steroids: Receptors and Function. New York, NY: Springer-Verlag.
View at Publisher | View at Google Scholar - Couse JF, Curtis SW, Washburn TF, et al. (1995). Analysis of transcription and estrogen insensitivity in the female mouse after targeted disruption of the estrogen receptor gene. Mol Endocrinol;9: 1441–1454.
View at Publisher | View at Google Scholar - Couse JF, Korach KS. (1999). Estrogen receptor null mice: what have we learned and what will they tell us? Endoc Rev; 20:358–417.
View at Publisher | View at Google Scholar - Fink G. (1988). Gonadotropin secretion and its control. In: Knobil E, Neil J, eds. The Physiology of Reproduction, vol. 1. New York, NY: Raven, 1349–1378.
View at Publisher | View at Google Scholar - George FW, Wilson JD. (1994). Sex determination and differentiation. In: Knobil E, Neil J, eds. The Physiology of Reproduction, vol. 1. New York, NY: Raven, 3–28
View at Publisher | View at Google Scholar - Green S, Chambon P. (1991). The estrogen receptor: from perception to the mechanism. In: Parker MG, ed. Nuclear Hormone Receptors: Molecular Mechanisms, Cellular Functions, Clinical Abnormalities. San Diego, CA: Academic, 15–38.
View at Publisher | View at Google Scholar - Isaacs JT. (1994). Role of androgens in prostate cancer. In: Litwack G, ed. Vitamins and Hormones, vol. 49. San Diego, CA: Academic, 433–502.
View at Publisher | View at Google Scholar - Jordan CV, ed. (1986). Estrogen/Antiestrogen Action and Breast Cancer Therapy. Madison, WI: University of Wisconsin Press.
View at Publisher | View at Google Scholar - Lindzey J, Kumar MV, Grossman M, Young C, Tindall, DJ. (1994). Molecular mechanisms of androgen action. In: Litwack G, Ed. Vitamins and Hormones, vol. 49. San Diego, CA: Academic, 383–432.
View at Publisher | View at Google Scholar - Luke MC, Coffey DS. (1994). The male sex accessory tissues: structure, androgen action, and physiology. In: Knobil E, Neil J eds. The Physiology. of Reproduction, vol. 1. New York, NY: Raven, 1435–1487.
View at Publisher | View at Google Scholar - Moudgil VK, Ed. (1985). Molecular Mechanism of Steroid Hormone Action: Recent Advances. New York, NY: Walter de Gruyter.
View at Publisher | View at Google Scholar - Parker MG, ed. (1995). Nuclear Hormone Receptors: Molecular Mechanisms, Cellular Functions, Clinical Abnormalities. San Diego, CA: Academic, 1991. Sluyser M. Mutations in the estrogen receptor gene. Hum Mutat; 6:97–103.
View at Publisher | View at Google Scholar - Smith EP, Boyd J, Frank GR, Takahashi H, Cophen RM, et al. (1994). Estrogen resistance caused by a mutation in the estrogen-receptor gene in a man. N Engl J Med; 331:1056–1061.
View at Publisher | View at Google Scholar - White PC. (1994). Genetic diseases of steroid metabolism. In: Litwack G, ed. Vitamins and Hormones, vol. 49. San Diego, CA: Academic, 131–195.
View at Publisher | View at Google Scholar
