

Review Article | DOI: https://doi.org/10.31579/2834-5010/028
Messenger RNA Based Vaccines and Their Immunological Effect on Diseases*
*Corresponding Author: Osama Mohammed Hasan, Dentistry College / Mustansiriyah University.
Citation: Osama M. Hasan, (2025), Messenger RNA Based Vaccines and Their Immunological Effect on Diseases, International Journal of Clinical Therapeutics, 4(1); DOI:10.31579/2834-5010/028
Copyright: © 2025, Osama Mohammed Hasan. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 02 January 2025 | Accepted: 09 January 2025 | Published: 17 January 2025
Keywords: mrna; transform cell; immunity; vaccine
Abstract
Because of its capabilities for fast development, potency, secure delivery, and promise for cost effective manufacture, mRNA vaccines are a promising vaccination technique. Many recent research has suggested that mRNA vaccines could be effective in treating a wide range of tumor and viral disorders where standard vaccine techniques have failed to stimulate protective immune responses. The inefficient and unstable in vivo distribution of mRNA has limited their application. Direct electroporation of mRNA vaccines into dendritic cells induced the generation of protective antibodies capable of destroying infected or transformed cells and inducing polyclonal CD8+ and CD4+ that mediated Ag specific T cell responses. in this review mRNA vaccines in detail were examined, as well as future objectives and challenges in the prevention of infectious diseases.
Introduction
Vaccine can protect against billion of microorganisms and can save hundreds of thousands of live each year [1]. Traditional vaccination methods, such as subunit vaccines and viruses that have been weakened and destroyed, provide long- term preservation against a different of deadly diseases [2].
Traditional vaccination approaches have been replaced by nucleic acid-based medicines, which have shown to be effective. In animals, data on the use of effective invitro transcribed (IVT) mRNA was recorded in (1990), and reporter mRNA were delivered to mice in a model, resulting in the discovery of protein expression. Core model of Watson and Crick state that DNA molecule is transcribed in to mRNA then eventually
translated to protein, which is the basis for nucleic acid vaccines. DNA or mRNAs penetration into a cell or organism results in the synthesis of a protein determined by nucleic acid sequences, which was found three decades ago. [3]. mRNA technology has a huge impact on vaccination research and development. Complex proteins that seem to be difficult to express using present methods may be capable to be produced [4].
Apart from DNA-based vaccines, mRNA has a number of benefits over subunit killed and weakened viruses. First, because there is no insertional mutagenesis process or infection danger, mRNA is a noninfectious and non-integrating platform. Furthermore, typical cellular mechanisms decay mRNA, and its invivo shelf - life can be controlled using many delivery techniques and changes. Second, mRNA is stabilized and translatable due to various changes. [5]. mRNA vaccines do not have the time-consuming production difficulties that afflict plasmid DNA, viral vectors, as well as other vaccines. Unlike plasmid DNA, mRNA does not require cell division to function and can be found for both mitotic cells and non-mitotic cell. In contrast to viral vector and plasmid, which have the ability to generate anti-DNA antibodies and autoimmunity by promoting long period of expression for specific target genes. mRNA is easier to regulate because it generates transitory creation of the encoded protein for a few days [6].
Production of mRNA vaccine
There are 2 type of mRNA vaccine currently in use, (1) non replicating vaccine and self-amplifying vaccine. Non replicating mRNA vaccine encode only the target antigen, (2) Self-amplifying mRNA vaccine on the other hand, also encode the virus's reproduction mode. This leads to prolonged and higher antigen expression, as well as greater vaccines stimulate the immune response. Self- amplifying and nonreplicating mRNA vaccine is utilized in infectious illness vaccines, while non-replicating mRNA has been used in tumor vaccine [7].
mRNA vaccines are made from plasmid using an RNA polymerase from a T3 phage, T7 phage, or Sp6 phage [8]. the mRNA construct has the same sequence, and the product should have an Open Reading Frame ORF flanked (UTRs) 5 caps, and (poly A) tail that encodes the target protein. The pDNA is linearized before being depraved by a DNase process to be the DNA-dependent RNA polymerase uses it as template for transcribed mRNA. The 5 cap and 3 poly(A) tail could be inserted during the invitro transcriptional stage or after transcription enzymatically [9]. In eukaryotic cells, the conventional 5 cap structure is an inverted 7 methyl guanosine m7G that is linked to the first nucleotides of the mRNA by a 5-5’ triphosphate bridges co-transcriptionally. The 5 cap's function is to boost the mRNA's translational efficacy and stability while simultaneously reducing its immunogenicity [10].
Because, poly (A tail) is very critical for mRNA stability & translation a typical poly (A) length that should be added to mRNA strand either with poly(A) polymerase or directly from the encoding DNA molecule [11]. the poly (A) tail length should be between 100 & 250 nucleotides. The cell type influences how long the poly (A) tail must be. The poly (A) tail promotes mRNAs efficiency and stability during translation. Modified adenosines can improve the poly (A) tail’s susceptibility to cellular RNasedigestion [12] Figure1
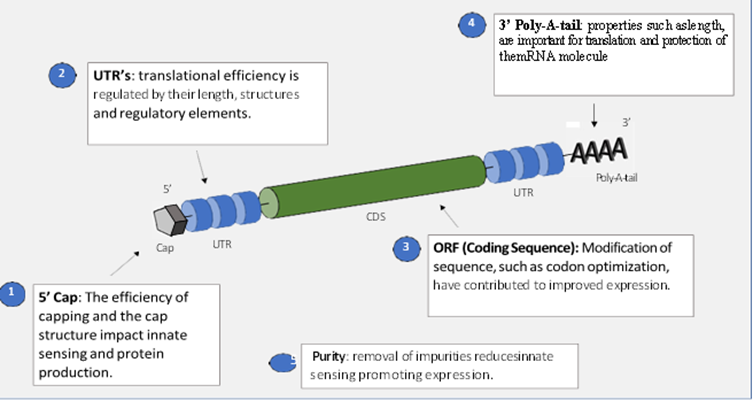
Figure 1: shown the performance of the mRNA construct [13]
The untransilation region (UTR) in sections (5′) & (3′) are critical for optimizing gene expression. The (3′) UTR length ,14 the structure of the 5′ UTR, and regulatory element in both UTR [13]. The protein of interest is encoded within the open reading frame. It can also have an impact on translation. Several codon optimization strategies have been developed with the goal of improving the translational process [14].
During IVT, modifying nucleotide can be utilized to stabilise IVT mRNA while reducing immunogenicity [15]. To name a few, adenosine with the N6 methyladenosine m6A and cytidine with the 95- methylcytidine -m5C also uridine with the 5-methyluridine m5U, 2- thiouridine s2U, or (pseudouridine) have all been found to decrease immunogenicity and induce translation activity of (IVT) mRNA [16].
Blocking cell signalling for protein kinase R (PKR) stimulation with modified nucleosides like N-1-methylpseudouridine or pseudouridine led to an improvement adaptive immune activation and antigen expression [17].
Methods of vaccine delivery based on mRNA.
In comparison to subunit, inactivated viral, live attenuated, and DNA vaccines, the use of an mRNA-based vaccination has some advantages. For starters, because mRNA is non integrating and noninfectious and there is no danger for infection and intigrated mutagenesis process. Also, mRNA is degenerate in normal cellular condition and it’s in vivo shelf life could be managed using various delivery techniques and modification systems [18].
In general, mRNA is produced in a cell- free environment using a DNA template and a transcription reaction. Because mRNA candidates must pass through membrane lipids to reach various nanoparticles such as Techniques for loading and delivering RNA into the cytoplasm have been devised, including protamines (LNPs) lipid polymer hybrid, golden nanoparticle, and cell-based delivery [19]. Among such lipid nanoparticles, LNPs have the most optimistic. They comprise phospholipids to comprise a lipid bilayer structures, cholesterol to keep the composition a lipid linked polyethylene glycol to extend the half-life of the compositions and very importantly, an ionizable cationic lipid to induce transfer of (mRNA) first from endosome to cytosol [20].
A cationic peptide called protamine has found to secure mRNA against serum Rnase digestion [21]. Protamine is a positive charge protein with a unique opportunity to interact with nucleic acids, such as mRNA, to enhance transfection and absorption. Positively charged protamine has been demonstrated to create a compound with mRNA by electrostatic interaction [22,23]. In order to increase mRNA uptake in vivo, physical methods of permeating the cell plasma membrane have been used. Early investigations suggested that a gene gun using a micro bullet technique may create messenger RNA connectd with gold nanoparticles in tissue [24]. Developing messenger RNA vaccines using electroporation-based cell- specific mRNA delivery is yet another option. By minimizing off-target effects, this can increase mRNA transport to target cells while lessening the required mRNA dose. The concept behind cell-specific messenger RNA delivery is that APCs (like dendritic cells) in these cell organs are in close proximity to T-lymphocytes, giving perfect conditions for functional enhancing T-cell stimulations in vivo [25]. Innate and adaptive immune responses to an mRNA-based vaccination.
The encoded proteins translated and delivered to the immuno system when the mRNA vaccination is administered. The natural process of a viral illness and the transactivation of the immune system's defense mechanisms is very similar to this technique. When exogenous mRNA reaches the cytoplasm, it is processed identically to endogenous mRNAs so foreign mRNA delivery to the cytosol is needed for Ag expression, however whether this is controlled by endosomal mRNAs is uncertain [26]. Tool like receptersTLR3 and TLR7/8 which recognize double stranded RNA and single stranded RNA, particularly, detect mRNA, as does the retinoic acid inducible gene I protein (RIG-I) pathway, which induces cell death and an inflammatory response. (RIG-I) is a cytosolic template matching receptors that stimulates type I interferon and the host immuno system by boosting type I interferon and identifying short double-stranded RNA [27]. mRNA spasticity may be useful and perhaps advantageous for vaccination since it may support adjuvant function to enhance dendritic cell DC maturation and thus stimulate robust B and T cell immuno responses in some cases. On other side innate immune detection of mRNA has been linked to antigen (Ag) expression inhibition, which may have a negative effect on the immune reaction [28]. In conjunction with treatment for cancer, dendritic cells might be infected with tumor-associated antigens (TAAs) encoding Mrna or total tumor RNA [29].. It has been shown that effective targeting of (IVT) Mrna nanoparticles causes robust effector reactions along memory T cells, and also IFN-mediated eradication of advanced cancers. Only the Ag presenting cell (APCs) can display peptides on both MHC-I and MHC-II, which are crucial to eliciting B cell and CD4+ T cell reactions, as well as antibody reactions. the development of adaptive immune responses in a timely and efficient manner the translated candidate Ag is transported to APCs through MHC I and MHC II subunits [30]. Without need of patient specific tumor derived cell or proteins Ags, DC transfected with mRNA encoding cancer related Ag can be used for immunization. [31]. Exterior messenger RNA is considered immune stimulatory because it stimulates innate immune cells such as through TLR (TLR8, TLR7, and TLR3) [32,33], and B cells are quickly stimulated by the (MYD88-TLR7- dependent - signaling pathways), providing activation for the upregulation of adaptive-immune response stimulated by mRNA vaccine [34]. Vaccine with two or even more mRNAs that express for a mixture of protein or lengthy peptide can enhance a polyclonal immuno response. HLA molecule constraints are eliminated, as is the chance of immunological escaping due to Ag loss, and using mRNAs coding for specific epitopes acquired from mutant proteins. In vaccinated individuals a significant antigen specific CD8+ T cell response is induced, and efficient CD4+ T cell-mediated cancer regression [35].
Conclusions
The nature of the mRNA vaccines technique in order for significant improvements in the optimization procedure with near limitless combinations of variants. This demands the application of translational sciences to induce formulation in order to be ready for further development and the selection of the best build. In the last two years, there's been an increase in preclinical research, which has been complemented by clinical experience. The rapid pace of research in mRNA vaccine will have not been achieved without major advancements in development of innate immuno detection of mRNA and invivo delivery mechanisms. Comprehensive mRNA research has allowed the translation of mRNA vaccines into clinical trials, leading in a startling level of exploitation in mRNA vaccines organizations.
References
- Pardi N, Hogan M.J, Porter F.W, Weissman D. mRNA vaccines-a new era in vaccinology. Nat. Rev. Drug Discov. 2018; 17: 261–279.
View at Publisher | View at Google Scholar - Plotkin, S.A. Vaccines: The fourth century. Clin. Vaccine Immunol. 2009; 16: 1709–1719.
View at Publisher | View at Google Scholar - Wolff J.A, Malone R.W, Williams P, Chong W, Acsadi G, Jani A, Agnes F, Philip L. Direct gene transfer into mouse muscle in vivo. Science 1990; 247: 1465–1468.
View at Publisher | View at Google Scholar - Cheng W. F. Enhancement of sindbis virus self-replicating RNA vaccine potency by linkage of herpes simplex virus type 1 VP22 protein to antigen. J. Virol. 2001 ;75: 2368–2376.
View at Publisher | View at Google Scholar - Thess, A. Sequence-engineered mRNA without chemical nucleoside modifications enables an effective protein therapy in large animals. Mol. Ther. 2015; 23 :1456–1464.
View at Publisher | View at Google Scholar - Hobernik D, Bros M. DNA vaccines- how far from clinical use. Int J Mol Sci. 2018;19 :3605.
View at Publisher | View at Google Scholar - Annkristin H, Stefan J, Peter Brossart. Clinical and immunological effects of mRNA vaccines in malignant diseases. Molecular Cancer. 2021; 20:52
View at Publisher | View at Google Scholar - Pardi N, Muramatsu H, Weissman D, Kariko, K. In vitro transcription of long RNA containing modified nucleosides. Methods Mol. Biol. 2013 ;969: 29–42.
View at Publisher | View at Google Scholar - Grudzien N. E. Synthesis of anti- reverse cap analogs (ARCAs) and their applications in mRNA translation and stability. Meth. Enzym. Ch. 2007;431: 203–227.
View at Publisher | View at Google Scholar - Hornung V, Ellegast J, Kim S, Brzózka K, Jung A, Kato H, et al. 5’- Triphosphate RNA is the ligand for RIG-I. Science. 2006; 314:994–7.
View at Publisher | View at Google Scholar - Gallie D. R. The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev.1991; 5: 2108–2116.
View at Publisher | View at Google Scholar - Strzelecka D, Smietanski M, Sikorski PJ, Warminski M, Kowalska J, Jemielity J. Phosphodiester modifications in mRNA poly(A) tail prevent deadenylation without compromising protein expression. RNA. 2020; 26:1815–1837
View at Publisher | View at Google Scholar - Leppek, K, Das R, Barna M. Functional 5ʹ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat. Rev. Mol. Cell Biol.2018 ;19: 158–174.
View at Publisher | View at Google Scholar - Mauro VP, Chappell SA. Considerations in the use of codon optimization for recombinant protein expression. In: Hacker DL, editor. Recombinant protein expression in mammalian cells. New York: Springer New York. 2018; 1850 :275–88
View at Publisher | View at Google Scholar - Hornung V, Ellegast J, Kim S, Brzózka K, Jung A, Kato H, Poeck H, Akira S, Conzelmann K.K, Schlee M, et al.50-Triphosphate RNA is the ligand for RIG-I. Science 2006; 314: 994–997.
View at Publisher | View at Google Scholar - Pardi N, Muramatsu H, Weissman D, Karikó K. In vitro transcription of long rna containing modified nucleosides. In: Rabinovich PM, editor. Synthetic messenger RNA and cell metabolism modulation. Totowa: Humana Press; 2013;969: 29–42.
View at Publisher | View at Google Scholar - Pardi, N.
View at Publisher | View at Google Scholar - Thess A, Grund S, Mui B.L, Hope M.J, Baumhof P, Fotin-Mleczek M, Schlake T. Sequence-engineered mRNA Without Chemical Nucleoside Modifications Enables an Effective Protein Therapy in Large Animals. Mol. Ther. 2015; 23: 1456–1464.
View at Publisher | View at Google Scholar - Islam M.A, Reesor E.K.G, Xu Y, Zope H.R, Zetter B.R, Shi J. Biomaterials for mRNA delivery. Biomater. Sci. 2015; 109: 78–87.
View at Publisher | View at Google Scholar - Pardi N, Hogan MJ, Weissman D. Recent advances in mRNA vaccine technology. Curr Opin Immunol. 2020; 65:14–20.
View at Publisher | View at Google Scholar - Hoerr, I., Obst, R., Rammensee, H. G. & Jung, G. In vivo application of RNA leads to induction of specific cytotoxic T lymphocytes and antibodies. Eur. J. Immunol. 2000; 30:1–7.
View at Publisher | View at Google Scholar - Scheel B, Teufel R, Probst J, Carralot J.P, Geginat J, Radsak M, Jaroslav D, Wagner H, Jung G, Rammensee H.G, et al. Toll-like receptor-dependent activation of several human blood cell types by protamine-condensed mRNA. Eur. J. Immunol. 2005; 35:1557–1566.
View at Publisher | View at Google Scholar - Barroso da Silva F.L, Derreumaux P,Pasquali
View at Publisher | View at Google Scholar - Qiu P, Ziegelhoffer P, Sun J, Yang, N. S. Gene gun delivery of mRNA in situ results in efficient transgene expression and genetic immunization. Gene Ther.1996 ;3: 262–268.
View at Publisher | View at Google Scholar - Kranz L.M, Diken M, Haas H, Kreiter S, Loquai C, Reuter K.C, Meng M, Fritz D Vascotto F, Hefesha, H. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016; 534:396–401.
View at Publisher | View at Google Scholar - Iavarone C, Ramsauer K, Kubarenko AV, Debasitis JC, Leykin I, Weber ANR, A point mutation in the amino terminus of TLR7 abolishes signaling without affecting ligand binding. J Immunol. 2011; 186:4213–22.
View at Publisher | View at Google Scholar - Kariko K, Buckstein M, Ni H, Weissman D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity 2005; 23: 165-75.
View at Publisher | View at Google Scholar - Weissman D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside modified, protein-encoding mRNA. Nucleic Acids Res. 2011;39: e142.
View at Publisher | View at Google Scholar - McNamara M.A, Nair S.K, Holl E.K. RNA-Based Vaccines in Cancer Immunotherapy. J. Immunol. Res. 2015; 2015:1- 9.
View at Publisher | View at Google Scholar - Kurts C, Robinson BWS, Knolle PA. Cross-priming in health and disease. Nat Rev Immunol. 2010; 10:403–14.
View at Publisher | View at Google Scholar - Su Z, Dannull J, Heiser A, Yancey D, Pruitt S, Madden J, Coleman D, Niedzwiecki D, Gilboa E, Vieweg J. Immunologicaland clinical responses in metastatic renal cancer patients vaccinated with tumor RNA- transfected
View at Publisher | View at Google Scholar - Chen N, Xia P, Li S, Zhang T, Wang TT, Zhu J. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life. 2017; 69:297–304.
View at Publisher | View at Google Scholar - Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of doublestranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 2001; 413:732–8.
View at Publisher | View at Google Scholar - Iavarone C, Ramsauer K, Kubarenko AV, Debasitis JC, Leykin I, Weber ANR, A point mutation in the amino terminus of TLR7 abolishes signaling without affecting ligand binding. J Immunol.
View at Publisher | View at Google Scholar
