

Research Article | DOI: https://doi.org/10.31579/2834-8087/005
Treatment of Male Albino Rats with 5-fluorouracil and/or Cyclophosphamide Caused Serious Alterations in Sexual Hormones, Histological Structure of the Testis, and Semen Quality
1 Pharmacology Department, Faculty of Medicine, Sabratha University, Libya.
2 Department of Environmental Studies, Institute of Graduate Studies and Research, Alexandria University.
3 Physiology Department, Faculty of Medicine, Sabratha University, Libya.
4 Department of Cell Biology, Medical Research Institute, Alexandria University.
*Corresponding Author: Azab Elsayed Azab, Physiology Department, Faculty of Medicine, Sabratha University, Libya.
Citation: Rabia A M Yahya, Ahmed M. Attia, Azab Elsayed Azab, Karema El.M.Shkal, and Mona A. Yehia (2022). Treatment of Male Albino Rats with 5-fluorouracil and/or Cyclophosphamide Caused Serious Alterations in Sexual Hormones, Histological Structure of the Testis, and Semen Quality. Archives of Clinical Investigation, 1(1) DOI:10.31579/2834-8087/005
Copyright: © 2022 Azab Elsayed Azab, This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received: 26 September 2022 | Accepted: 03 October 2022 | Published: 10 October 2022
Keywords: cyclophosphamide; 5-fluorouracil; sexual hormones; semen quality; histological structure of the testis; male albino rats
Abstract
Background: The therapeutic effect of Cyclophosphamide (CPA) is thus attributed to phosphoramide mustard and acrolein that leads to the formation of high level of reactive oxygen species (ROS), which results in decreased antioxidant activity. Excessive production of ROS could also culminate in oxidative stress.
Objectives: The aim of this study is to evaluate the effect of sub lethal dose of the cyclophosphamide, 5-FU combination of 5-FU and CPA on testicular function, and histopathological alterations in male albino rats. These functions were monitored through some selected biochemical parameters and confirmed by DNA, RNA concentration and histopathological examination.
Materials and Methods: Twenty-eight male adult rats were grouped randomly into four groups (n=7 each group), Five for biochemical studies and 2 for histological studies. Group I (control): Rats were injected with saline intraperitoneally and at a dose of 1.0 ml/kg b.w. for 14 days. Group II cyclophosphamide (CPA): Cyclophosphamide at a dose of 10 mg/kg day by day through i.p. to rats for 14 days. Group III Fluorouracil (5-FU): 5-Fluorouracil at a dose of 10 mg/kg day by day in saline was given through i.p. to rats for 14 days. Group IV (CPA+5-FU): Rats were given CPA followed by 5-FU at a dose of 10 mg/kg per day (day by day) through i.p. to rats for 14 days. At the end of the experimental period, rats were anesthetized using light ether. Blood and testes tissue samples were taken and prepared for biochemical and histological measurements. Biochemical parameters in rat serum and tissues were evaluated.
Results: Treatment of male rats with CPA, 5-FU and their combination caused a significant decrease in sperm count, motility while increased dead and abnormal sperms compared to those of control. Co-treatment of CPA and 5-FU caused a significant decrease in sperm count, sperm motility and increase in abnormal sperms compared to individual treatment of CPA and 5-FU. Levels of testosterone, luteinizing hormone (LH) and follicle stimulating hormone (FSH) were decreased in the CPA, 5-FU and their combination-treated group. Co-treatment of 5-FU and CPA synergistically reduced testicular hormones compared with individual chemotherapy. Cyclophosphamide, 5-FU and their combination decreased testis DNA and RNA concentrations compared to the control. The combination of CPA and 5-FU was more effective to DNA and RNA compared to individual administration of the chemotherapy. Epididymis of the control rats showed normal histological structure with normal sperm density. Large tubules lined by pseudostratified columnar epithelium with abundant stereocilia projection from their surface. Long and dense processes of sperm tails were observed within the Lumina. The epididymis tubules of CPA-exposed rats were smaller and the majorities of them were free from mature spermatozoa. Epithelium contained numerous numbers of sloughed cells with cell debris in Lumina. 5-FU treated rat showed atrophy as well as necrosis, the arrangement of germ cells was disordered. The number of cell layers of the seminiferous tubules was significantly reduced. Epididymis of CPA+5-FU-treared rats showed necrotic degeneration and atrophy of the germinal cells, decrease spermatid when compared to CPA.
Conclusion: It could be concluded that treatment of mammals with chemotherapy is associated with the production of free radicals that lead to hazardous alterations in sexual hormones, DNA, and RNA concentration, and histopathological structure in tistes and semin quality of male albino rats. However, 5-FU and CPA combination could produce a serious alteration in these parameters. Future work should consider combined chemotherapy regimens. Toxicological studies must be performed before using drugs as combination before application. Further research is required on toxicological impacts of drugs and pollutants mixtures.
Introduction
Cyclophosphamide (CP) is one of the most invasive chemotherapeutic agents. Clinical evidence showed toxic side effects of CP in multiple organ systems [1,2]. reported that administration of 65 mg/kg i.p.CPA to Wistar rats for 5 days induced reproductive toxicity as evidenced by significantly lowered levels of FSH, LH and testosterone, perturbation of sperm characterization. Histopathological examination of the testis showed ballooned seminiferous tubules with loosed connective tissues and vacuolation of testicular interstitium. [1] reported that Serum testosterone concentration was significantly (p<0>) decreased in adult male Wistar rats treated i.p. with 6.1 mg/kg of CP /day for 50 days. The male patients treated with cyclophosphamide have demonstrated disturbed gonadotrophin secretion associated with testicular damage along with low blood level of testosterone [3].
5-FU is used to treat colorectal cancer as well as other kinds of cancer. Its cytotoxic effects on cancer cells exert through the inhibition of thymidylate synthase and the incorporation of its metabolites into RNA and DNA [4, 5]. 5-FU is an analogue of uracil with a fluorine atom at the C-5 position in place of hydrogen. It rapidly enters the cell using the same facilitated transport mechanism as uracil [6]. 5-FU is converted intracellularly to several active metabolites: fluorodeoxyuridine monophosphate (FdUMP), fluorodeoxyuridine triphosphate (FdUTP) and fluorouridine triphosphate (FUTP) [7]. [5] concluded that a small dose of 5-FU affect the patient fertility by disturbing the testes histology and sperm morphology.
Objectives
The aim of this study is to evaluate the effect of sub lethal dose of the cyclophosphamide, 5-FU combination of 5-FU and CPA on testicular function, and histopathological alterations in male albino rats. These functions were monitored through some selected biochemical parameters and confirmed by DNA, RNA concentration and histopathological examination.
2. Objectives
The aim of this study is to evaluate the effect of sub lethal dose of the cyclophosphamide, 5-FU combination of 5-FU and CPA on testicular function, and histopathological alterations in male albino rats. These functions were monitored through some selected biochemical parameters and confirmed by DNA, RNA concentration and histopathological examination.
3. Materials and Methods
The present research was conducted in the Environmental Toxicology Laboratory, Department of Environmental Studies, Institute of Graduate Studies and Research, Alexandria University, Egypt.
3.1. Chemicals
Reduced glutathione (GSH), 1-chloro-2,4-dinitrobenzene, thiobarbituric acid and all other chemicals were purchased from Sigma Chemical Company (Saint Louis, USA). Other chemicals from Sigma or other trusted commercial sources. Cyclophosphamide and 5-Fluorouacil were purchased from
3.2. Animals
Twenty eight male adult rats (Sprague Dawley) with average body weight of 180±10 g were obtained from Faculty of agriculture, Alexandria, and acclimatized for two weeks prior to the experiment. They were assigned to four groups and housed in Universal galvanized wire cages at room temperature (22-25°C) and in photoperiod of 12h/day. Animals were provided with balanced commercial diet.
3.3. Experiential protocol
Twenty eight male adult were grouped randomly into fo
ur groups (n=7 each group).
- Group I (control): Rats were injected with saline intraperitoneally and at a dose of 1.0 ml/kg b.w. for 14 days.
- Group III cyclophosphamide (CPA): Cyclophosphamide at a dose of 10 mg/kg day by day through i.p. to rats for 14 days [8].
- Group III Fluorouracil (5-FU): 5-Fluorouracil at a dose of 10 mg/kg day by day [9] in saline was given through i.p. to rats for 14 days. .
- Group IV (CPA+5-FU): Rats were given CPA followed by 5-FU at a dose of 10 mg/kg per day (day by day) through i.p. to rats for 14 days.
- At the end of the experimental period, rats were anesthetized using light ether. Blood samples were taken from the vena cava of rat heart. Tubes were used to compile blood drawn from the heart directly; serum formation, blood was allowed to set for 30 min at 4oC to clot, then centrifuged for 5 minutes at 1000 xg. Packed cells were discarded and the supernatant serum samples were decanted and stored into capped sterile polyethylene tubes at -20oC until used (within 24 hours). The abdominal cavity of each rat was opened where the testes were excised. Tissue was blotted on a filter paper to remove excess buffer, and the tissue was weighed prior the addition of 5-10 ml cold 50 mM potassium phosphate buffer, pH 7.5 containing 1 mM EDTA per gram tissue. Then, tissue was homogenized using a glass pestle (glass homogenizer). The homogenate was centrifuged at 10,000 x g for 15 min at 4°C, and the supernatant was collected and stored at -80 oC for further use.
3.7. Testicular RNA and DNA concentration
The RNA concentration in the samples was determined by precipitation of the nucleic acids in 0.5 M HClO4, after which the RNA was hydrolysed ice by incubation in 0.3 M KOH at 60°C for 1 h. After preremoval of DNA by 0.5 M HClO4 precipitation, the RNA concentration was determined by boiling the samples for 30 min in 6 M HCl, 0.01
4. Results
4.1. Serum levels of testosterone, luteinizing hormone, and follicle stimulating hormone
A significant decrease was shown in serum levels of testosterone, luteinizing hormone (LH) and follicle stimulating hormone (FSH) in the CPA, 5-FU and their combination-treated group. Co-treatment of 5-FU and CPA synergistically reduced testicular hormones compared with individual chemotherapy (Tables .1 and Figures 1-3).
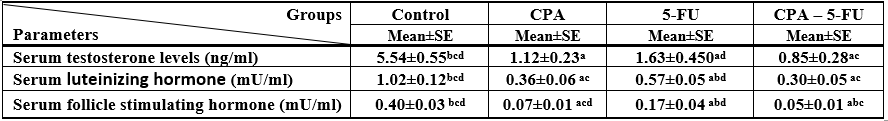
Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil, a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
Table 1: Serum testosterone, luteinizing hormone, and follicle stimulating hormone levels of rat treated with cyclophosphamide and/or 5-fluorouracil
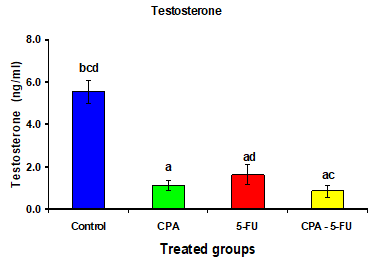
Figure 1: Serum testosterone (ng/ml) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups;
b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
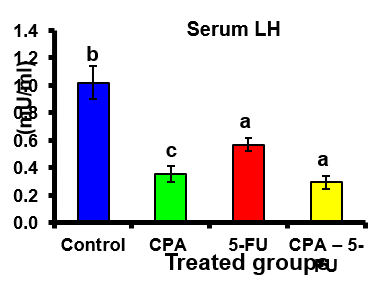
Figure 2: Serum Luteinizing hormone (LH) (mU/ml) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
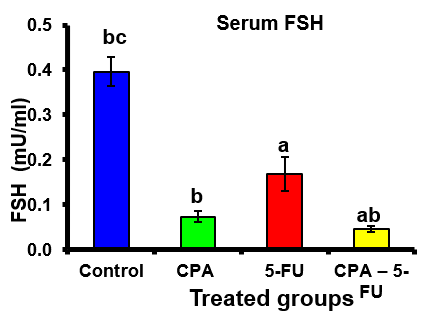
Figure 3: Serum follicle stimulating hormone (FSH) (mU/ml) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
4.5. Testes DNA and RNA
Cyclophosphamide, 5-FU and their combination decreased (P<0>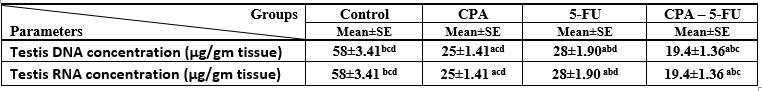
Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil, a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
Table 2: Serum testosterone, luteinizing hormone, and follicle stimulating hormone levels of rat treated with cyclophosphamide and/or 5-fluorouracil
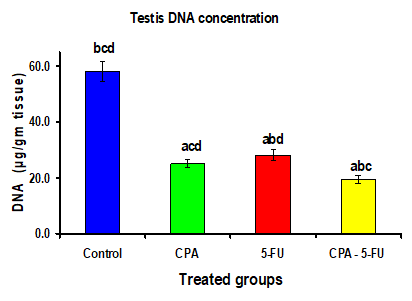
Figure 4: Testis DNA concentration (µg/gm tissue) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
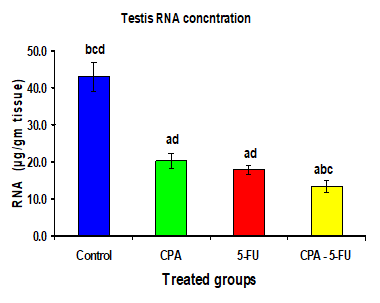
Figure 5: Testis RNA concentration (µg/gm tissue) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
4.6. Histopathological findings
Epididymis of the control rats showed normal histological structure with normal sperm density. Large tubules lined by pseudostratified columnar epithelium with abundant stereocilia projection from their surface. Long and dense processes of sperm tails were observed within the Lumina (Figures .6). The epididymis tubules of CPA-exposed rats were smaller and the majorities of them were free from mature spermatozoa. Epithelium contained numerous numbers of sloughed cells with cell debris in Lumina (Figure. 7). Photomicrographs of cauda epididymis of 5-FU treated rat showing atrophy as well as necrosis, the arrangement of germ cells was disordered. The number of cell layers of the seminiferous tubules was significantly reduced (Figure .8). Epididymis of CPA+5-FU-treared rats showed necrotic degeneration and atrophy of the germinal cells, decrease spermatid when compared to CPA (Figure .9).
Figure 6: Photomicrographs of cauda epididymis of control rat showing normal section in rat testis with normal lyeding cells, spermatocytes, seminiferous epithelium, spermatid. Uniform seminiferous tubules lined by normal layers of spermatogenic cells up to mature sperm formation
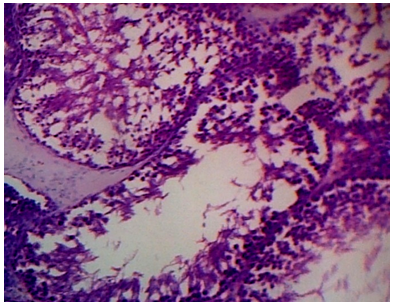
Figure 7: Photomicrographs of cauda epididymis of CPA treated rat showing severe testicular degeneration. Note that sperm density is severely decreased. Some tubules are empty and others have few spermatozoa and LC variable sized and shaped seminiferous tubules lined by few layers of spermatogenic cells with wide lumina
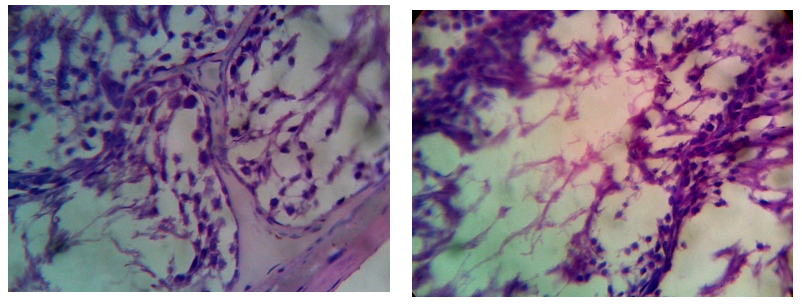
Figure 8: Photomicrographs of cauda epididymis of 5-FU treated rat showing atrophy as well as necrosis, the arrangement of germ cells was disordered. The number of cell layers of the seminiferous tubules was significantly reduced.
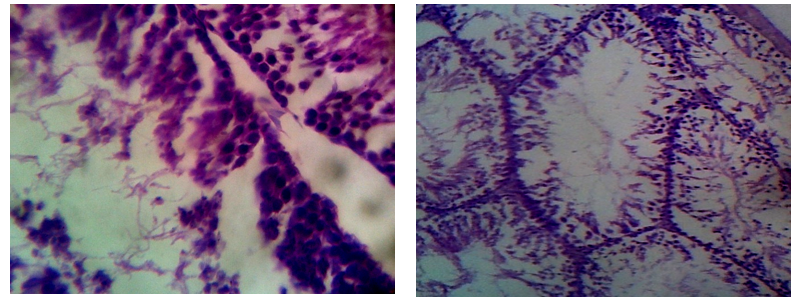
Figure 9: Photomicrographs of cauda epididymis of combined treated rat with 5-FU and CPA showing necrotic degeneration and atrophy of the germinal cells, decrease spermatid.
4.3. Sperm characteristics
Treatment of male rats with CPA, 5-FU and their combination caused a significant decrease (p < 0>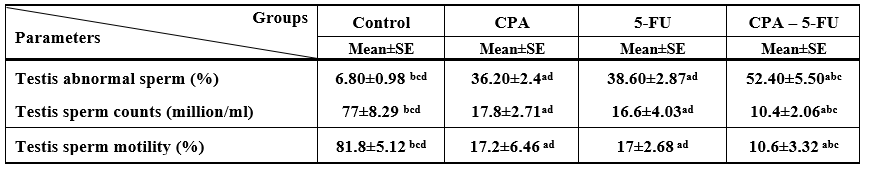
Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil, a Comparison of control and other groups; b Comparison of CPA
and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
Table 3: Testis abnormal sperm, sperm counts, and sperm motility of rat treated with cyclophosphamide and/or 5-fluorouracil
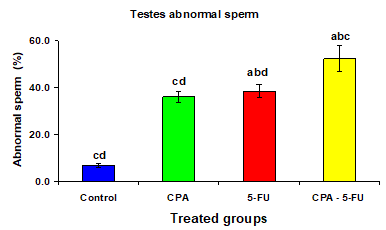
Figure 10: Testis abnormal sperm (%) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
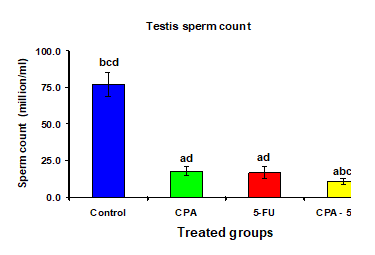
Figure 11: Testis sperm count (million/ml) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups.

Figure 12: Testis sperm motility (%) of rat treated with cyclophosphamide, fluorouracil and their combination. Significance at P > 0.05. CPA: cyclophosphamide; 5-FU: Fluorouracil. a Comparison of control and other groups; b Comparison of CPA and other groups; c Comparison of 5-FU and other groups; d Comparison of CPA – 5-FU and other groups
5. Discussion
A significant decrease was shown in serum levels of testosterone, luteinizing hormone (LH) and follicle stimulating hormone (FSH) in the CPA, 5-FU and their combination-treated group. Co-treatment of 5-FU and CPA synergistically reduced testicular hormones compared with individual chemotherapy. Testosterone is produced by leydig cell in the testes under the influence of LH. Intratesticular testosterone is an absolute prerequisite for normal spermatogenesis. The decrease in the mean testosterone levels of 5-FU and CPA treated rat served as proof for the damage of testis. These results demonstrate that tested chemotherapy caused temporary interference of normal male reproductive system with treatment.
The results revealed that CPA, 5-FU and their combination reduced serum testosterone level, the endogenous antioxidant enzyme activities as well as the non-enzymatic scavenger (GSH) in treated rats. It also suppressed lipid peroxidation and significantly enhanced the lowered TAC and most of the decreased testicular marker enzyme activities. In accordance with previous work [16], CPA administration induced a state of oxidative stress in the rat testis, demonstrated by enhanced lipid peroxidation and compromised antioxidant defense system revealed by decreased TAC and GSH levels with concomitant reduction in GR activities. Oxidative stress is the result of an imbalance between ROS generation and the intracellular capacity for removing ROS, subsequently leading to excessive damage in the cell [17]. Increase in testicular ROS level was previously reported in CPA-treated rats [18]. Spermatozoa are particularly susceptible to ROS-induced injury and peroxidative damage because of their high concentrations of polyunsaturated fatty acids and low antioxidant capacity [19].
Cylophosphamide is an alkylating agent commonly used in combination cancer chemotherapy regimens. It is also used alone as an immunosuppressive agent for conditions such as glomerulonephritis [20] and Bechcet's disease [21]. There are a number of reports on the effects of this drug on fertility. Adult patients treated daily with cyclophosphamide (1-2 ng/kg) for more than 4 month showed oligozoospermia or azoospermia [20-22]. This change in spermatogenic function was associated with increases in serum follicle-stimulating hormone (FSH) levels [21]. The return of fertility was unpredictable but in some cases it took a number of years [23].
It is well known that utilization of oxygen represents an efficient mechanism for aerobic organisms to generate energy, but ROS are also produced within the biological systems [24]. Under normal conditions to protect cells against oxidative damage, the potential harmful effects of ROS and free radicals are effectively eliminated by the antioxidant defense systems such as antioxidant enzymes and non-enzymatic factors [25]. Normally, cells possess a well-developed biochemical defense system, comprising low-molecular weight free radical scavengers, i.e. glutathione (GSH), vitamin C, vitamin E and complex enzymes, including GPx, SOD and CAT [26]. When the balance between the generation and elimination of ROS is broken, as a result of these events, biomacromolecules including DNA, membrane lipids and proteins are damaged by ROS-mediated oxidative stress [27, 28].
[29] had reported that CPA requires microsomal oxidation in the liver to yield its active metabolite, 4-hydroxy-CPA in target cells that spontaneously decomposes to phosphoramid mustard which exerts cytotoxic effect by the induction of DNA single strand breaks as well as crosslinks which result in different types of damage. Also, CPA has been shown to induce apoptosis in the target cells through the modulation of signaling through caspases, Bcl2, Bax, NF-Kappa B, and MAP kinases [30, 31]. [32] had reported that CPA involves inhibition of cell division due to cross-linking of the drug to DNA.
Results of both biochemical and histological investigations showed that CPA, 5-FU and their combination induced a marked reproductive toxicity through induction of oxidative stress. In this study, testicular reduction in testosterone, LH and FSH levels were occurred due to CPA, 5-FU and their combination treatment. Furthermore CPA, 5-FU treatments induced a marked decrease of spermatogenesis, as confirmed by our histological findings of sperm density reduction in epididymis. In explanation to the present finding, it has been reported that antineoplastic agents can disturb Leydig cells directly [33].
Thus, the reduction in circulating testosterone is supposed to be resulting from a direct poisonous effect of CPA and 5-FU on the Leydig cells. Steroidogenesis in the male rats is stimulated by hypothalamic gonadotropin releasing hormone (GnRH) effecting to induce the production and release of LH, which binds to LH receptors on the membrane of Leydig cells to upregulate testosterone production [34]. The reduction in LH level perhaps is result of damage in the negative feedback control of hypothalamic-pituitary axis [35]. In addition, it is likely that the dysfunction of the pituitary in LH releasing was resulted from damage to the cell membrane-mediated signaling mechanisms involved in releasing LH to the blood.
In the present study, epididymal sperm count decreased by CPA treatment while the number of dead and abnormal sperms increased and motility did not show significant difference. [36] showed that the incidence of male infertility following CPA chemotherapy resulted from changes in sperm parameters. Germ cells in testes are vulnerable to DOX-induced oxidative stress [37]. Anthracyclines like DOX exert their antitumor properties as well as other organ toxicity by intracellular producing of free radicals and ROS accompanied by intercalation with DNA and consequent inhibition of topoisomerase [38].
This increased oxidative stress effects on the sperm membranes, proteins and DNA [39-41]. Therefore, DNA damage may be liable for the increased level of abnormal spermatozoa forms. As confirmed by acridine orange staining, treatment with DOX causes single/double strand breaks in sperm DNA. In the case of the adult rat testis, the gonadotoxic drug DOX induces programmed cell death in meiotic spermatocytes and type A and intermediate spermatogonia by intercalating into DNA to create strand breaks and by preventing topoisomerase II activity [42]. These genotoxic alterations up-regulate expression of p53, an essential mediator of cell cycle stop considered to inhibit DNA replication in the presence of DNA damage, resulting apoptosis and finally drop in sperm counts [42].
In response to some anticancer drugs like anthracyclines, the number of male germ cells undergoing apoptosis increases several folds. Because the most sensitive cells to CPA and 5-FU are the early spermatogenic cells and primary spermatocytes, treatment with chemotherapy may lead to the loss of proliferating immature germ cells and finally of mature spermatozoa [43]. Therefore, a possible reason for the disruption of spermiogenesis in the CYT-treated rats is failure of testosterone dependent attachment of spermatids to Sertoli cells [44].
In a normal status, secretion of GnRH from hypothalamus stimulating the anterior pituitary gland to produce FSH and LH which in turn bring about gonadal hormones secretions (testosterone and estrogen) in testes. Reduced hypothalamic sensitivity to negative feedback effects of androgens increases GnRH secretion and consequently the amount of gonadotropin and androgen secretion increases which cause testicular growth and the incidence of other secondary sex characteristics. GnRH analogues with continuous stimulation of GnRH secretion, inhibits the decrement in secretion of LH and FSH hormones [45]. A GnRH analogue increases the secretion of LH and FSH in first injection, but thereafter, despite the presence of GnRH, gonadotropin secretion decreases [46]. Gonadotropin receptors are located on the testis and seminiferous tubules, decreased spermatogenesis which is caused by the chemotherapy agent disrupts the pituitary gonadal axis. In the rat chemotherapy the level of FSH is controlled based on spermatogenesis. Because the inhibition of meiosis by the chemotherapy agent, the spermatogenesis phase is quite impossible to observe [47, 48]. In the present study the levels of FSH and more distinctly LH are reduced, consequently the spermatogenesis negatively affected by chemotherapy. According to Brinkworth [49] cyclophosphamide-induced genetic damage to cells causes sexual harm.
According to Masta et al. [50] cross connections in the active parts of chromosomes are damage in chemotherapy. Evidences show that, the damages to the DNA of the male sex cells caused by chemicals and drugs lead to gene mutations and consequently congenital malformation which are transferable [49]. Men treated with anticancer drugs are more likely to experience permanent infertility and defects in the gonads [51]. Since the male sex cell division rate is very high, thus it is sensitive to anti-cancer or antimitotic agents. The results of this study confirmed that tissue damages in testicular tissue are higher in cyclophosphamide and 5-FU treated group than in the group treated with 5-FU plus CPA. The reason for reduction in the effect of combined chemotherapy is due to antagonist action leads to increase in non-enzymatic and enzymatic antioxidant and reduction in oxidative damage.
Testicular toxicity by chemotherapeutic agents is considered as inevitable side effect of cancer treatment in male patients. A strategy to diminish the side effects of anticancer drugs with preservation of chemotherapeutic efficacy is necessary. The present study reconfirmed the testicular toxicity of cyclophosphamide.
Cyclophosphamide has alkylating properties that result in nucleotide base mispairs and DNA/DNA or DNA/protein cross-linking that lead to major disruptions in nucleic acid function and the inhibition of DNA synthesis [52]. Cyclophosphamide undergoes biotransformation by hepatic microsomal cytochrome P450 isoenzymes to aldophosphamide, which is spontaneously broken down to produce phosphoramide mustard and acrolein. Phosphoramide mustard is responsible for anti-cancer effects, while acrolein is associated with the toxic effects observed during CPA therapy [53]. As CPA is an alkylating agent that causes more damage into rapidly dividing cells, the drug is expected to affect the seminiferous epithelium and reduce the number of spermatozoa produced. The number of spermatozoa in the cauda epididymis provides a good estimate of spermatozoa reserves.
Cyclophosphamide treatment has also been shown to cause oxidative stress [54]. Mammalian spermatozoa are rich in polyunsaturated fatty acids and thus very susceptible to ROS attack [55]. Excessive ROS increases germ cell apoptosis and detrimental effect on spermatozoa resulting in decrease in sperm viability, sperm motility and increase in morphology defects with deleterious effects on sperm capacitation and acrosome defects. Lipid peroxidation (LPO) destroys the structure of lipid matrix in the membranes of spermatozoa, and it is associated with loss of motility and defects of membrane integrity [56].
Therefore stress could play an important role in the induction of sperm abnormalities, namely higher susceptibility of sperm DNA to denature and fragment [57]. Cyclophosphamide and 5-FU treated rats in the present study had a significant increase in testes LPO and a significant drop in antioxidant levels suggesting the presence of free-radical toxic stress within the testes. Lipid peroxidation serves as an index of intensity of oxidative stress
6. Conclusion
It could be concluded that treatment of mammals with chemotherapy is associated with the production of free radicals that lead to hazardous alterations in sexual hormones, DNA, and RNA concentration, and histopathological changes in tistes and semin quality of male albino rats. However, 5-FU and CPA combination could produce a serious alteration in these parameters. Future work should consider combined chemotherapy regimens. Toxicological studies must be performed before using drugs as combination before application. Further research is required on toxicological impacts of drugs and pollutants mixtures.
References
- Hosseini, A., Zare, S., Borzouei, Z., & Pakdel, F. G. (2018). Cyclophosphamide-induced testicular toxicity ameliorate by American ginseng treatment: An experimental study. International Journal of Reproductive BioMedicine, 16(11), 711.
View at Publisher | View at Google Scholar - Ayoka, O.A., Ojo, O.E., Imafidon, E.C., Ademoye, K.A. and Oladele, A.A. (2016). Neuro-endocrine effects of aqueous extract of Amaranthus viridis (Linn.) leaf in male Wistar rat model of cyclophosamide -induced reproductive toxicity. Toxicology Reports, 3: 608-619.
View at Publisher | View at Google Scholar - Hoorweg-Nijman, J.J., Delemarre-vande-Wall, H.A., DeWall, F.C., Behrendt, H., (199)2. Cyclophosphamide-induced disturbance of gonadotropin secretion manifesting testicular damage. Acta Endocrinol. 126, 143–148.
View at Publisher | View at Google Scholar - Longley, D. B., Harkin, D. P. and Johnston, P. G. (2003) 5- fluorouracil: mechanisms of action and clinical strategies. Nat. Rev. Cancer. 3: 330–338.
View at Publisher | View at Google Scholar - Gad El-Hak, H. N., Moawad, T. I. S., & Hafez, G. A. A. (2017). Effects of 5-fluorouracil on testes histology and sperm morphology assay in mice. Egyptian Academic Journal of Biological Sciences, D. Histology & Histochemistry, 9(1), 1-9.
View at Publisher | View at Google Scholar - Wohlhueter, R. M., McIvor, R. S. and Plagemann, P. G. (1980). Facilitated transport of uracil and 5-fluorouracil, and permeation of orotic acid into cultured mammalian cells. J. Cell Physiol., 104: 309–319.
View at Publisher | View at Google Scholar - Diasio, R. B. and Harris, B. E. (1989). Clinical pharmacology of 5-fluorouracil. Clin. Pharmacokinet. 16: 215–237.
View at Publisher | View at Google Scholar - Muralikrishnan G, Amalan Stanley V, Sadasivan Pillai K (2001). Dual role of vitamin C on lipid profile and combined application of cyclophosphamide, methotrexate and 5-fluorouracil treatment in fibrosarcoma-bearing rats. Cancer Lett 169:115–120.
View at Publisher | View at Google Scholar - Subramaniam S, Shyamala Devi CS (1995) Vitamin E protects intestinal basolateral membrane from CMF-induced damages in rat. Indian J Physiol Pharmacol 39:263–266
View at Publisher | View at Google Scholar - Chirgwin, J.M., Przybyla, A.E., Macdonald, J. and Rutter, W.J. (1979). Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18: 5294–5299.
View at Publisher | View at Google Scholar - Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989). Molecular cloning: a laboratory manual, 2nd edn. New York: Cold Spring Harbor Laboratory
View at Publisher | View at Google Scholar - Baker, F.J.; Silverton, R.E. Introduction to Medical Laboratory Technology, 6th ed.; Butter Worth: London, UK, 1985.
View at Publisher | View at Google Scholar - Narayana, K., D’Souza, U.J.A. and Rao, K.P.S. (2005). Effect of ribavirin on epididymal sperm count in rat. Indian J. Physiol. Pharmacol., 46: 97–101.
View at Publisher | View at Google Scholar - Narayana, K., D’souza, U.J.A. and Rao, K.P.S. (2002). Ribavirin induced sperm shape abnormalities in Wistar rats. Mutation research, 5(13): 193-96.
View at Publisher | View at Google Scholar - Howell, D.C. (1995). Fundamental statistics for the behavioral sciences, 3rd ed.. Duxbury press. An imprint of Wads Worth publishing company Belmont. California. pp. 163-166.
View at Publisher | View at Google Scholar - Motawi, T.M.K., Sadik, N.A.H. and Refaat, A. (2010). Cytoprotective effects of DL-alpha-lipoic acid or squalene on cyclophosphamide-induced oxidative injury: an experimental study on rat myocardium, testicles and urinary bladder., Food Chem. Toxicol. 48: 2326–2336.
View at Publisher | View at Google Scholar - Vladimir-Knežević, S., Blažeković, B., Kindl, M., Vladić, J., Lower-Nedza, A.D. and Brantner, A.H. (2014). Acetylcholinesterase inhibitory, antioxidant and phytochemical properties of selected medicinal plants of the Lamiaceae family., Molecules, 19: 767–782.
View at Publisher | View at Google Scholar - Selvakumar, E., Prahalathan, C., Sudharsan, P.T. and Varalakshmi, P. (2006). Protective effect of lipoic acid on cyclophosphamide-induced testicular toxicity., Clin. Chim. Acta., 367: 114–119.
View at Publisher | View at Google Scholar - Vernet, P., Aitken, R.J. and Drevet, J.R. (2004). Antioxidant strategies in the epididymis., Mol. Cell. Endocrinol., 216:31–39.
View at Publisher | View at Google Scholar - Fairley, K.F., Barrie, J.U. and Johnson, W. (1972). Sterility and testicular atrophy related to cyclophosamide therapy. Lancet, 1:568-569.
View at Publisher | View at Google Scholar - Fukutani, K., Ishida, H., Shinohara, M., Minowada, S., Niijima, T., Hijikata, K. and Izawa, Y. (1981). Suppression of spermatogenesis in patients with Bechet's disease treated with cyclophosamide and colchicine. Fertil Steril., 36: 76-80.
View at Publisher | View at Google Scholar - Qureshi, M.S.A., Pennington, J.H., Goldsmith, H.J. and Cox, P.E. (1972). Cyclophosamide therapy and sterility. Lancet 2: 1290-1291.
View at Publisher | View at Google Scholar - Buchanan, J.D., Fairley, K.F., Barrie, J.U. (1975). Return of spermatogenesis after stopping cyclophosamide therapy. Lancet 2: 156-157.
View at Publisher | View at Google Scholar - Cutler, R.G. (1991) Human longevity and aging: possible role of reactive oxygen species. Ann. N Y Acad. Sci., 621:1–28.
View at Publisher | View at Google Scholar - Hu, Y., Rosen, D.J. and Zhou, Y. (2005). Mitochondrial manganesesuperoxide dismutase expression in ovarian cancer: role in cell proliferation and response to oxidative stress. J. Biol. Chem., 280:39485–39492.
View at Publisher | View at Google Scholar - Yao, P., Li, K., Song, F., Zhou, S., Sun, X., Zhang, X., Nussler, A.K. and Liu, L. (2007). Heme oxygenase-1 upregulated by Ginkgo biloba extract: potential protection against ethanol-induced oxidative liver damage. Food Chem. Toxicol., 45:1333–1342.
View at Publisher | View at Google Scholar - Tian, L., Cai, Q. and Wei, H. (1998). Alterations of antioxidant enzymes and oxidative damage to macromolecules in different organs of rats during aging. Free Radic. Bio. Med., 24:1477–1484.
View at Publisher | View at Google Scholar - Qian, Z.J., Jung, W.K. and Kim, S.K. (2008). Free radical scavenging activity of a novel antioxidant peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana shaw. Bioresource Technol., 99:1690–1698
View at Publisher | View at Google Scholar - Matalon, S. T.; Ornoy, A.; Lishner, M. (2004): Review of the potential effects of three commonly used antineoplastic and immunosuppressive drugs (cyclophosphamide, azathioprine, doxorubicin on the embryo and placenta). Reproductive Toxicol., 18: 219 – 230.
View at Publisher | View at Google Scholar - Mirkes, P. E.; Wilson, K. L.; Cornel, L. M. (2000): Teratogen-induced activation of ERK, JNK and P38 MAP kinases in early post implantation murine embryos. Teratol., 62: 1425.
View at Publisher | View at Google Scholar - Torchinsky, A.; Lishanski, L.; Wolstein, O.; Shepshelovich, J.; Orenstein, H.; Savion, S.; Zaslavsky, Z.; Carp. H.; Brill, A.; Dinstein, R.; Toder, V.; Fein, A. (2002): N F Kappa B DNA-binding activity in embryos responding to a teratogen Cyclophosphamide. BMC Dev. Biol., 2: 2.
View at Publisher | View at Google Scholar - Bosanquet, A. G.; Sturm, I.; Weider, T.; Essmann, F.; Bosanquet, M. I.; Head, D. J.; Dorken, B.; Daniel, P. T. (2002): Bax expression correlates with cellular drug sensitivity to doxorubicin, cyclophosphamide and chlorambucil but not to fludrabin, cladribin or cortico-steroids in B cell chronic lymphocytic leukemia. Leukemia J., 16: 1035.
View at Publisher | View at Google Scholar - Ait Hamadouche, N., Slimani, M., Merad-Boudia, B., Zaoui, C. (2009). Reproductive Toxicity of Lead Acetate in Adult Male Rats. Am. J. Sci Res., 3: 38-50.
View at Publisher | View at Google Scholar - Gamal Hozayen, W. (2012). Effect Of Hesperidin and Rutin On Doxorubicin Induced Testicular Toxicity in Male Rats. Int J Food Nutr Sci., 1: 31-42.
View at Publisher | View at Google Scholar - Kovacs, M., Schally, A.V., Nagy, A., Koppan, M., Groot, K. (1997). Recovery of pituitary function after treatment with a targeted cytotoxic analog of luteinizing hormone- releasing hormone. Proc Natl Acad Sci., 94: 1420-1425.
View at Publisher | View at Google Scholar - Howell, S.J. and Shalet, S.M. (2001). Testicular function following chemotherapy. Hum Reprod., 7: 363-369.
View at Publisher | View at Google Scholar - Hrdina, R., Gersl, V., Klimtova, I., Simunek, T., Mach, J. and Adamcova, M. (2000). Anthracycline-induced cardiotoxicity. Acta Medica., 43: 75-82.
View at Publisher | View at Google Scholar - Ichihara, S., Yamada, Y., Kawai, Y., Osawa, T., Furuhashi, K., Duan, Z., and Ichihara, G.. (2007). Roles of oxidative stress and Akt signaling in doxorubicin cardiotoxicity. Biochem Biophys Res. Com., 359: 27-33.
View at Publisher | View at Google Scholar - Kalender, Y., Yel, M. and Kalender, S. (2005). Doxorubicin hepatotoxicity and hepatic free radical metabolism in rats. The effects of vitamin E and catechin. Toxicology, 209: 39-45.
View at Publisher | View at Google Scholar - Saalu, L.C., Osinubi, A.A., Oguntola, J.A., Adeneye, I.O., Benebo, A.S. (2010). The Delayed Testicular Morphologic Effects of Doxorubicin and the Rejuvinating Role of Grapefruit Seed Extract. Int. J. Pharm., 6: 192-199.
View at Publisher | View at Google Scholar - Favier, A.E. (1992). The role of zinc in reproduction, Hormonal mechanisms. Biol Trace Elem Res., 32: 363-382.
View at Publisher | View at Google Scholar - Hou, M.I., Chrysis, D., Nurmio, M., Parvinen, M., Eksborg, S., Soder, O., Jahnukainen, K. (2005). Doxorubicin Induces Apoptosis in Germ Line Stem Cells in the Immature Rat Testis and Amifostine Cannot Protect against This Cytotoxicity. Cancer Res., 65: 9999-10005.
View at Publisher | View at Google Scholar - Zanetti, S.R., Maldonado, E.N. and Aveldaño, M.I. (2007). Doxorubicin Affects Testicular Lipids with Long-Chain (C18-C22) and Very Long-Chain (C24-C32) Polyunsaturated Fatty Acids. Cancer Res., 67: 6973-6980.
View at Publisher | View at Google Scholar - Rezvanfar, M.A., Sadrkhanlou, R.A., Ahmadi, A., Shojaei-Sadee, H., Rezvanfar, M.A., Mohammadirad, A., Salehnia, A. and Abdollahi, M. (2008). Protection of cyclophosphamide-induced toxicity in reproductive tract histology, sperm characteristics, and DNA damage by an herbal source; evidence for role of free-radical toxic stress. Hum Exper Tox., 27: 901-910.
View at Publisher | View at Google Scholar - Hellerstedt, B.A. and Pienta, K.J. (2002). The current state of hormonal therapy for prostate cancer. CA Cancer J Clin. 52:154–179.
View at Publisher | View at Google Scholar - Brawer, M.K. (2001). The evolution of hormonal therapy for prostatic carcinoma. Rev Urol. 3:S1–S9.
View at Publisher | View at Google Scholar - Tohda, A., Matsumiya, K., Tadokoro, Y., Yomogida, K., Miyagawa, Y., Dohmae, K., et al. (2001). Testosterone suppresses spermatogenesis in juvenile spermatogonial depletion (jsd) mice. Biol Reprod. 65:532–537.
View at Publisher | View at Google Scholar - Shetty, G., Weng, C.C., Meachem, S.J., Bolden-Tiller, O.U., Zhang, Z., Pakarinen, P., et al. (2006). Both testosterone and FSH independently inhibit spermatogonial differentiation in irradiated rats. Endocrinology. 147:472–482.
View at Publisher | View at Google Scholar - Brinkworth, M.H. (2000). Paternal transmission of genetic damage: findinds in animals and humans. Int J Androl. 23:123–135.
View at Publisher | View at Google Scholar - Masta, A, Gray, PJ, Philips, DR. (1994). Molecular basis of nitrogen mustard effects on transcription process: role of depurination. Nucleic Acids Res. 1994;22:3880–3886.
View at Publisher | View at Google Scholar - Roeser, H.P., Stocks, A.E. and Smith, A.J. (1978). Testicular damage due to cytotoxic drugs and recovery after cessation of therapy. Aust NZ J Med. 8:250–254.
View at Publisher | View at Google Scholar - Crook, T. R., Souhami, R. L., McLean, A. E. (1986). Cytotoxicity, DNA cross-linking, and single strand breaks induced by activated cyclophosphamide and acrolein in human leukemia cells. Cancer Res., 46: 5029–5034.
View at Publisher | View at Google Scholar - LeBlanc, G.A. and Waxman, D.J. (1990). Mechanisms of Cyclophosphamide Action on Hepatic P-450 Expression. Cancer Res. 50(9): 5720-5726.
View at Publisher | View at Google Scholar - Oh, M.S., Chang, M.S., Park, W., Kim, D.R., Bae, H., Huh, Y. and Park, S.K. (2007). Yukmijihwang-tang protects against cyclophosphamide-induced reproductive toxicity. Reproductive Toxicology 24: 365–370.
View at Publisher | View at Google Scholar - Aitken, R.J. and Roman, S.D. (2008). Antioxidant systems and oxidative stress in the testes. Oxidative Medicine and Cellular Longevity: 1(1): 15-24.
View at Publisher | View at Google Scholar - Sanocka, D. and Kurpisz, M. (2004). Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology: l2:12.
View at Publisher | View at Google Scholar - Shalaby, M.A. and El Zorba, H.Y. (2010). Protective effect of celery oil, Vitamin E and their combination against testicular toxicity in male rats. Global veterinaria: 5(2): 122-128.
View at Publisher | View at Google Scholar
